Niche construction is the process by which organisms alter environmental states, thereby modifying the conditions that they, and other organisms, experience, and the sources of natural selection in their environments.
Organisms adapt to their environments through natural selection. However, they also modify natural selection through niche construction. In this way they influence evolution.
For examples of niche construction, watch or download the slideshow and visit our resources page.
Emperor penguins are the only vertebrate species able to breed during the Antarctic winter. They huddle together to try to keep warm in an icy landscape that can be as cold as 50 degrees Celsius below zero. But how warm is it inside the huddle?
By huddling together for just two hours, penguin bodies can raise the temperature in the middle of the huddle to as high as 37 degrees above zero! The penguins are able to construct a warm environment, even in a blizzard (Gilbert et al, 2007)!
Despite living on land for millions of years, earthworms have retained the physiology of the freshwater species from which they evolved. How can they survive with such ‘poor’ morphological adaptation?
Through niche construction. Earthworms process soil in ways allowing them to draw water into their bodies more effectively, creating a simulated aquatic environment on land (Turner, 2000). It is as if they built their own swimming pools!
Hermit crabs use shells as safe homes to live in. They modify these shells to increase their inner size and reduce their weight. As they grow, they need larger shells. But the only shells big enough are those carried by other hermit crabs. What do they do?
Hermit crabs gather, waiting for other hermit crabs to change shells, and then jump into a suitable vacated one. Niche construction creates a biological market, leading to the evolution of sociality in a hitherto solitary species (Laidre, 2012).
Female dung beetles manufacture and bury a brood ball of dung and insert into it a faecal pedestal onto which they lay an egg. Through niche construction they provide a safe home, food, and microbiome for their developing young (Schwab et al 2016).
Developing larvae also processes the brood ball, changing microbiome composition. Experiments show that maternal and offspring niche construction strongly affect offspring size, fitness and trait characteristics (Schwab et al 2016, 2017).
Some trees increase the rate of forest fires through shedding flammable needles, cones, seeds and oils onto the ground. This is an adaptation that benefits the trees but damages competitors. How has this affected their evolution?
The trees have evolved fire resistance, rapid resprouting after fire, and often require fire to reproduce (Schwilk 2003). Forest fires benefit them, but not the competition.
Even traits such as the timing of germination and flowering can be niche-constructing traits. How?
The timing of germination and flowering determine the seasonal environment experienced by plants and their offspring. This alters many traits, the expression of genetic variation of those traits, and natural selection on those traits (Donohue, 2013).
Humans are champion niche constructors. Through agriculture, deforestation, urbanization, control of fire, and the domestication of plants and animals, humans have modified environments in dramatic ways, even on a global scale.
The domestication of cattle and consumption of dairy products is a compelling example of human niche construction. Dairy culture generated natural selection favouring genes allowing adults to metabolise lactose in milk (Gerbault et al, 2011).
These snails (Euchondrus spp.) feed on lichens that grow on and under rocks in the Negev desert. Each snail is very small, and has a tiny effect on its environment, and yet their activities have a massive effect on the entire desert ecosystem. How?
The snails feeding breaks up rocks and inadvertently generates tonnes of soil. This soil supports a plant community, which in turn supports insects and birds. The snails play a vital role in constructing the desert ecosystems.
The yeast Saccharomyces cerevisiae facilitates its own propagation by modifying fruit to produce chemicals that attract Drosophila, and then hitches a ride (Buser et al 2014). Here niche construction drives coevolutionary interactions.
Experimental studies with bacteria demonstrate that niche construction evolves rapidly, under a broad range of conditions (Callahan et al 2014).
Niche construction can occur even without a physical change in the environment. Through migrating, birds transform their local environment from a harsh location with little food or mating opportunities, to one rich with food and mates.
Waterpepper smartweeds alter the shapes of their leaves when their light conditions are different, thereby altering the amount of light they receive. This is known as ‘experiential niche construction’ (Sultan 2015).
More desert plants have small leaves or spines, to conserve water. So how can this plant, the desert rhubarb (Rheum palaestinum), survive with huge leaves?
It’s a self-irrigating plant! Its leaves channel water towards its taproot, which grows and then shrinks again, producing chemical exudates that line the cavity, where water gathers (Lev-Yadun et al, 2009).
Traditionally, niche construction has been viewed as a source of environmental change, but not a cause of evolution. Evolutionary processes are generally thought of as phenomena that directly change gene frequencies.
A broader view of evolutionary causation recognizes that niche construction biases the direction of evolution by systematically modifying selection.
Previous
Next
Emperor penguins are the only vertebrate species able to breed during the Antarctic winter. They huddle together to try to keep warm in an icy landscape that can be as cold as 50 degrees Celsius below zero. But how warm is it inside the huddle?
By huddling together for just two hours, penguin bodies can raise the temperature in the middle of the huddle to as high as 37 degrees above zero! The penguins are able to construct a warm environment, even in a blizzard (Gilbert et al, 2007)!
Despite living on land for millions of years, earthworms have retained the physiology of the freshwater species from which they evolved. How can they survive with such ‘poor’ morphological adaptation?
Through niche construction. Earthworms process soil in ways allowing them to draw water into their bodies more effectively, creating a simulated aquatic environment on land (Turner, 2000). It is as if they built their own swimming pools!
Hermit crabs use shells as safe homes to live in. They modify these shells to increase their inner size and reduce their weight. As they grow, they need larger shells. But the only shells big enough are those carried by other hermit crabs. What do they do?
Hermit crabs gather, waiting for other hermit crabs to change shells, and then jump into a suitable vacated one. Niche construction creates a biological market, leading to the evolution of sociality in a hitherto solitary species (Laidre, 2012).
Female dung beetles manufacture and bury a brood ball of dung and insert into it a faecal pedestal onto which they lay an egg. Through niche construction they provide a safe home, food, and microbiome for their developing young (Schwab et al 2016).
Developing larvae also processes the brood ball, changing microbiome composition. Experiments show that maternal and offspring niche construction strongly affect offspring size, fitness and trait characteristics (Schwab et al 2016, 2017).
Some trees increase the rate of forest fires through shedding flammable needles, cones, seeds and oils onto the ground. This is an adaptation that benefits the trees but damages competitors. How has this affected their evolution?
The trees have evolved fire resistance, rapid resprouting after fire, and often require fire to reproduce (Schwilk 2003). Forest fires benefit them, but not the competition.
Even traits such as the timing of germination and flowering can be niche-constructing traits. How?
The timing of germination and flowering determine the seasonal environment experienced by plants and their offspring. This alters many traits, the expression of genetic variation of those traits, and natural selection on those traits (Donohue, 2013).
Humans are champion niche constructors. Through agriculture, deforestation, urbanization, control of fire, and the domestication of plants and animals, humans have modified environments in dramatic ways, even on a global scale.
The domestication of cattle and consumption of dairy products is a compelling example of human niche construction. Dairy culture generated natural selection favouring genes allowing adults to metabolise lactose in milk (Gerbault et al, 2011).
These snails (Euchondrus spp.) feed on lichens that grow on and under rocks in the Negev desert. Each snail is very small, and has a tiny effect on its environment, and yet their activities have a massive effect on the entire desert ecosystem. How?
The snails feeding breaks up rocks and inadvertently generates tonnes of soil. This soil supports a plant community, which in turn supports insects and birds. The snails play a vital role in constructing the desert ecosystems.
The yeast Saccharomyces cerevisiae facilitates its own propagation by modifying fruit to produce chemicals that attract Drosophila, and then hitches a ride (Buser et al 2014). Here niche construction drives coevolutionary interactions.
Experimental studies with bacteria demonstrate that niche construction evolves rapidly, under a broad range of conditions (Callahan et al 2014).
Niche construction can occur even without a physical change in the environment. Through migrating, birds transform their local environment from a harsh location with little food or mating opportunities, to one rich with food and mates.
Waterpepper smartweeds alter the shapes of their leaves when their light conditions are different, thereby altering the amount of light they receive. This is known as ‘experiential niche construction’ (Sultan 2015).
More desert plants have small leaves or spines, to conserve water. So how can this plant, the desert rhubarb (Rheum palaestinum), survive with huge leaves?
It’s a self-irrigating plant! Its leaves channel water towards its taproot, which grows and then shrinks again, producing chemical exudates that line the cavity, where water gathers (Lev-Yadun et al, 2009).
Traditionally, niche construction has been viewed as a source of environmental change, but not a cause of evolution. Evolutionary processes are generally thought of as phenomena that directly change gene frequencies.
A broader view of evolutionary causation recognizes that niche construction biases the direction of evolution by systematically modifying selection.
The niche construction perspective
The niche construction perspective was originally introduced into evolutionary biology through the pioneering writings of Conrad Waddington and Richard Lewontin. The perspective is encapsulated in the below quote:
The organism influences its own evolution, by being both the object of natural selection and the creator of the conditions of that selection.
However, niche construction is not restricted to the biological adaptations of organisms, but also stems from their developmental plasticity, their by-products, and their acquired characters. Organisms are viewed as active agents that impose order on the conditions of their existence, and direction on their development and evolution.
Lewontin RC. 1983. Gene, organism, and environment. In: Bendall DS, ed. pp. 273-285. Cambridge: Cambridge University Press This accessible essay introduces the niche construction perspective and critiques conventional evolutionary approaches to adaptation.
Lewontin RC. 2000. The Triple Helix: Gene, Organism, and Environment. Cambridge: Harvard University Press Another readable introduction to the niche-construction perspective by the pioneer of this way of thinking.
Odling-Smee FJ, Laland KN, Feldman MW. 1996. Niche construction.American Naturalist. 147:641–648 A brief, but non-technical, introduction to niche construction for evolutionary biologists.
Odling-Smee FJ, Laland KN, Feldman MW 2003. Niche Construction: The Neglected Process in Evolution. Princeton: Princeton University Press An authoritative, rigorous and extensive introduction to niche construction theory.
In this blog post, Kevin Laland explains why niche construction is defined broadly.
Contents
Operational definition of niche construction
The following criteria test for the presence of niche construction (Criteria 1 and 2) and determine when niche construction affects evolution (Criterion 3) (Matthews et al, 2014):
Criterion 1) An organism (i.e., a candidate niche constructor) must significantly modify environmental conditions.
Criterion 2) The organism-mediated environmental modifications must influence selection pressures on a recipient of niche construction.
Criterion 3) There must be a detectable evolutionary response in a recipient of niche construction that is caused by the environmental modification of the niche constructor.
Odling-Smee FJ, Laland KN, Feldman MW 2003. Niche Construction: The Neglected Process in Evolution. Princeton: Princeton University Press An authoritative, rigorous and extensive introduction to niche construction theory.
Matthews B, De Meester L, Jones CG, Iberlings BW, Bouma TJ, Nuutinen V, van der Koppel J & Odling-Smee J. 2014. Under niche construction: an operational bridge between ecology, evolution and ecosystem science.Ecological Monographs. 84.2: 245–263. This article clarifies how niche construction and ecosystem engineering can be distinguished, and illustrates how the former can be investigated.
Perturbational and relocational niche construction
The defining characteristic of niche construction is not organism-driven modification of the environment per se (a special case known as perturbational niche construction), but rather modification of the organism’s niche (that is, modification of the relationship between the organism and its environment). Hence the term ‘niche construction’ includes such cases as dispersal, migration and habitat selection, where organisms relocate in space to modify both the environments that they leave and that they enter (known as relocational niche construction).
Inceptive and counteractive niche construction
If an environmental factor is already changing, or has changed, organisms may oppose or cancel out that change, a process labelled counteractive niche construction. For instance, many wasps and bees will engage in temperature regulation of their nests, heating it up through muscular activity in the cold, and placing droplets of water on the surface, allowing it to cool through evaporation, in the heat. Counteractive niche construction is therefore conservative or stabilizing, and it functions to buffer organisms from shifts in factors away from states to which they have been adapted.
Experiential niche construction
Organisms can also make adjustments that change their experience of the environment, without changing the environment itself (experiential niche construction) (Sultan 2015, Chiu 2019). For instance, penguins huddle to keep warm, whilst plants shift the orientation of their leaves to maximize input from the sun .
Odling-Smee FJ, Laland KN, Feldman MW 2003. Niche Construction: The Neglected Process in Evolution. Princeton: Princeton University Press An authoritative, rigorous and extensive introduction to niche construction theory.
Timeline of events central to the development of Niche Construction Theory
The scientific study of niche construction dates back to the early writings on ecological succession of William King in the 17th century. There were important 19th century contributions by Charles Darwin (1851) on corals and earthworms (1881), Lewis Henry Morgan (1868) on the beaver, Nathaniel Shaler’s (1892) analyses of how animals affect soils, and Frederic Clements’ (1916) classic work on plant ecological succession.
Charles Darwin
More recently, Niche Construction Theory was brought to prominence through the foundational 20th century work of developmental biologist Conrad Waddington and evolutionary biologists Richard Lewontin and John Odling-Smee. Also influential was the work of Erwin Schrödinger.
Schrödinger and Waddington’s contributions
Nobel prize winning physicist Erwin Schrödinger’s (1944) work already implied that niche construction is universal.
Erwin Schrödinger
Schrödinger pointed out that living organisms are far-from-equilibrium systems relative to their physical surroundings. To preserve their out-of-equilibrium status, while creating order (structure, organization) in their bodies and their immediate surrounds, Schrödinger emphasized that organisms must actively do work on their environment. In other words, living organisms can only survive by constantly engaging in niche construction. This activity cannot be random: organisms must change environments in systematic, directional ways.
Waddington’s view of the causes of evolution, with the exploitive system highlighted. Adapted from Fig. 1 in Waddington (1959).
He wrote (1959, p1635-6):
Animals … are usually surrounded by a much wider range of environmental conditions than they are willing to inhabit. They live in a highly heterogeneous ‘ambience’, from which they themselves select the particular habitat in which their life will be passed. Thus the animal by its behaviour contributes in a most important way to determining the nature and intensity of the selective pressures which will be exerted on it.
Richard Lewontin’s contribution
Harvard evolutionary biologist Richard Lewontin (1982, 1983, 2000) brought niche construction theory reasoning to prominence within the evolutionary biology community.
His writings stressed how genes, organisms and environments are in reciprocal interaction with each other in such a way that each is both cause and effect (Lewontin, 1983, p281):
“Organisms do not adapt to their environments: they construct them out of the bits and pieces of their external world.”
According to Lewontin, this construction process has a number of features:
(1) Organisms determine what is relevant. While stones are part of a thrush’s environment, tree bark is part of a woodpecker’s, and the underside of leaves part of a warbler’s. It is the life activities of these birds that determine which parts of the world, physically accessible to all of them, are actually parts of their environments….
(2) Organisms alter the external world as it becomes part of their environments. All organisms consume resources by taking up minerals, by eating. But they may also create the resources for their own consumption, as when ants make fungus farms, or trees spread out leaves to catch sunlight….
(3) Organisms transduce the physical signals of the external world. Changes in external temperature are not perceived by my liver as thermal changes but as alterations in the concentrations of certain hormones and ions….
(4) Organisms create a statistical pattern of environment different from the pattern in the external world. Organisms, by their life activities, can damp oscillations, for example in food supply by storage, or in temperature by changing their orientation or moving. They can, on the contrary, magnify differences by using small changes in abundance of food types as a cue for switching search images. They can also integrate and differentiate. Plants may flower only when a sufficient number of days above a certain temperature have been accumulated.
Lewontin’s stance can be contrasted with the more traditional view, expressed by fellow evolutionary biologist George Williams (1992):
Adaptation is always asymmetrical; organisms adapt to their environment, never vice versa.
However, while Lewontin’s articles focused attention on niche construction, they were generally interpreted as a critique of adaptationism, and it was not apparent to most biologists how these ideas could be translated into practical science.
Together with Kevin Laland and Marcus Feldman, Odling-Smee began the first systematic investigation of niche construction. The team documented and categorized the extent of niche construction in living organisms, devised methods by which its ecological and evolutionary consequences could be investigated, and derived dedicated theory to explore these questions (Odling-Smee et al. 1996, 2003; Laland et al. 1996, 1999).
Additional attempts to model niche construction, and explore its ecological and evolutionary ramifications, followed (e.g. Lehmann 2008; Kylafis & Loreau 2008; van Dyken & Wade 2012). Over the next decade research into niche construction increased rapidly, with a large number of experimental and theoretical studies across a broad range of fields. Several influential academic books make niche construction a central concept.
a display of books related to niche construction
In 2015, evolutionary ecologist Sonia Sultan published Organism & Environment: Ecological Development, Niche Construction, and Adaptation. Sultan documents extensive evidence for niche construction and its evolutionary consequences, across a wide variety of taxa, and develops the concept of niche construction in important respects.
Waddington CH. 1959 Evolutionary systems – animal and human. Nature 183:1634-1638. Waddington was ahead of his time in recognizing the importance of niche construction (which he called the ‘exploitative system’) for the evolutionary process.
Lewontin R. 1983. Gene, organism, and environment. In Evolution: From Molecules to Men, ed DS Bendall, pp. 273–85. Cambridge: Cambridge University Press. This accessible essay introduces the niche construction perspective and critiques conventional evolutionary approaches to adaptation.
Odling-Smee, F.J. Niche constructing phenotypes. In. H.C. Plotkin, ed., The Role of Behavior in Evolution, 73-132. Cambridge: MIT Press. Here Odling-Smee first makes the argument that niche construction should be recognized as an evolutionary process, and introduces the concept of ‘ecological inheritance.’
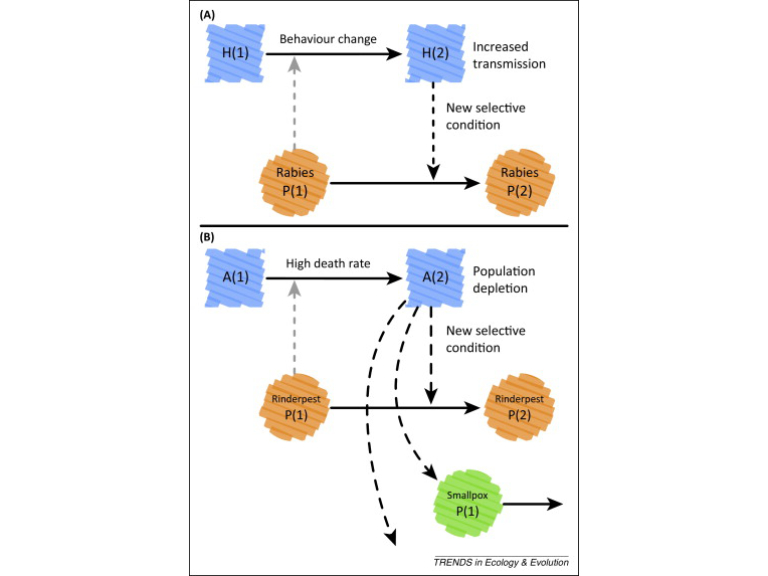
Niche construction overlaps with several other concepts deployed within ecology and evolution (Odling-Smee et al, 2003). Three of these are particularly noteworthy: ecosystem engineering, eco-evolutionary dynamics, and the extended phenotype.
Ecosystem Engineering
Niche construction and ecosystem engineering are related ideas that arose independently in separate fields (niche construction in evolutionary biology, and ecosystem engineering in ecology), but which both place emphasis on changes that organisms make in environments.
Ecosystem engineers are organisms that modify their physical surroundings (e.g., light environment, physical habitat structure) so as to modulate the availability of resources or energy fluxes in an ecosystem (Jones et al. 1994, 1997). By comparison, niche constructors are organisms that alter selection pressures of a recipient organism by modifying any aspect of the abiotic and biotic environment. While evidence of ecosystem engineering would not constitute evidence of niche construction, nonetheless ecosystem engineers are excellent candidates for being niche constructors, particularly when they generate persistent ecosystem effects (Matthews et al, 2014).
In recent times, the two fields have been regarded as complementary and mutually reinforcing, with much cross-citation, and joint publications (e.g. Odling-Smee et al, 2013; Matthews et al, 2014).
From Fig 1. of Matthews et al. 2014
Eco-evolutionary dynamics
Eco-evolutionary dynamics has a very broad focus that includes both the ecological and evolutionary responses of populations to interactions between organisms and their environments (Fussmann et al. 2007, Post & Palkovacs 2009). Evolution by niche construction is closely related to eco-evolutionary dynamics but the two concepts have different emphases. Eco-evolutionary dynamics must include at least two organisms and at least one evolutionary and one ecological effect, whilst neither of these conditions is necessary for evolution by niche construction (Matthews et al 2014). This means there are cases of evolution by niche construction that do not constitute eco-evolutionary dynamics.
Matthews et al (2014) emphasize that while some of the potential mechanisms of niche construction are well-studied, others are not, and that niche construction has value in drawing attention to these gaps. Several research areas in ecology and evolution could be expanded to test for new examples of evolution by niche construction by measuring evolutionary responses of organism-mediated environmental modifications.
The extended phenotype
Extended phenotypes are adaptations expressed outside of the body of the constructor, such as birds’ nests or termites’ mounds (Dawkins1982). ‘Extended phenotype’ is a narrower term than niche construction, because it is restricted to forms of environmental modification that are biological adaptations.
In contrast, niche construction theory emphasizes that niche construction also incorporates evolutionary by-products that trigger evolutionary episodes. This is vital, since in ecosystems the ecological and evolutionary consequences that flow from niche-constructing by-products are likely to be as substantial as those flowing from niche-constructing adaptations.
Extended phenotypes also exclude aspects of niche construction that depend on acquired knowledge, for instance, human cultural processes. This is an important limitation for scholars of anthropogenic change, since very little human niche construction can accurately be characterized as a biological adaptation.
Finally, the environmental modifications produced by niche-constructing organisms, such as beaver’s dams or termite mounds, may persist through repetition and reconstruction for longer than the individual constructors, and may continue to modulate the impact of these effects on subsequent generations of the same or other populations, a legacy described as an ‘ecological inheritance’.
Theoretical models have shown that niche-constructing by-products, cultural niche construction and ecological inheritance can all strongly affects evolutionary dynamics (Laland et al 1999, 2001; Lehmann 2007, 2008; Creanza & Feldman, 2014).
Further distinctions between niche construction and the extended phenotype are discussed here.
Conceptual change
Over-and-above the aforementioned distinctions, niche construction theory differs from the study of ecosystem engineering, eco-evolutionary dynamics, and the extended phenotype in suggesting that niche construction is an evolutionary process.
Niche construction theory emphasizes how niche construction can scale up across individuals in populations and over time to generate stable and directional modification of environmental components. As a result, niche construction generates consistent and sustained forms of natural selection, and thereby imposes biases on evolution (Laland et al, 2015).
Niche construction theory can be viewed as part of a wider movement that embraces a broader conception of evolutionary causation. For further discussion, see Niche construction and the EES.
Waddington CH. 1959 Evolutionary systems – animal and human. Nature 183:1634-1638. Waddington was ahead of his time in recognizing the importance of niche construction (which he called the ‘exploitative system’) for the evolutionary process.
Lewontin R. 1983. Gene, organism, and environment. In Evolution: From Molecules to Men, ed DS Bendall, pp. 273–85. Cambridge: Cambridge University Press. This accessible essay introduces the niche construction perspective and critiques conventional evolutionary approaches to adaptation.
Odling-Smee, F.J. Niche constructing phenotypes. In. H.C. Plotkin, ed., The Role of Behavior in Evolution, 73-132. Cambridge: MIT Press. Here Odling-Smee first makes the argument that niche construction should be recognized as an evolutionary process, and introduces the concept of ‘ecological inheritance.’
Niche construction theory contrasts with conventional conceptualizations of evolution in recognizing niche construction as an evolutionary cause or process. The niche-construction perspective seeks to explain the adaptive complementarity of organism and environment in terms of a dynamic, reciprocal interaction between the processes of natural selection and niche construction.
The classic niche constructor
The distinctive aspects of the niche-construction perspective are illustrated by the familiar example of the beaver. Generally, the evolutionary consequences of beaver dam building are modeled in terms of fitness ‘payoffs’ to the underlying genes; selection favors dam-building alleles over their alternatives. According to Richard Dawkins (1982), who exemplifies the traditional standpoint, dams can be viewed as ‘extended phenotypes’Biological adaptations expressed outside of the body of the organism. – adaptations expressed outside of the constructor’s body, but that evolve in a manner no different from other adaptations.
From the niche-construction perspective this stance is unsatisfactory, both because it misses part of the causal story and because it discourages consideration of additional forms of selective feedback. When a beaver builds a dam, creating a lake and influencing river flow, it not only affects the propagation of dam-building genes but dramatically changes its local environment, affecting nutrient cycling, decomposition dynamics, the structure of the riparian zone, and plant and community composition and diversity. It follows that beaver dam building must also transform selection acting on a host of other beaver traits. The active agencyThe capacity of living organisms to act on, and in, their world, and to modify their experience of it, including in ways that are neither predetermined, nor random. of beavers in constructing these modified selection pressures and thereby acting as co-directors of their own evolution (not to mention that of other species) currently goes unrecognized.
Often the modifications produced by niche-constructing organisms persist for longer than the individual constructors, continuing to modulate the impact of environment on subsequent generations, a legacy described as an ‘ecological inheritance’The inheritance, via an external environment, of one or more natural selection pressures previously modified by niche-constructing organisms.. Descendants of the dam builder will experience modified selection so long as the dam, lake, and transformed environment remain. Given that dams are frequently maintained by families of beavers for decades, that could be considerably longer than the lifetime of an individual beaver. In addition to genes, offspring inherit a modified selective environment.
While it is possible to study this feedback using established evolutionary theory, for instance through models of habitat selection, co-evolution or maternal effects, nonetheless the magnitude and significance of niche construction remains underappreciated, in part because standard evolutionary theory does not encourage attention to such phenomena.
Conversely by treating niche construction as an evolutionary process – one that is influenced by, but not solely reducible to, earlier natural selection – niche construction theory generates novel hypotheses and new methods that stimulate research. The perspective is rapidly growing, precisely because researchers from multiple disciplines find it useful.
Contents
Evolutionary significance of niche construction
Several factors render niche construction of evolutionary significance:
(1) Niche construction biases natural selection
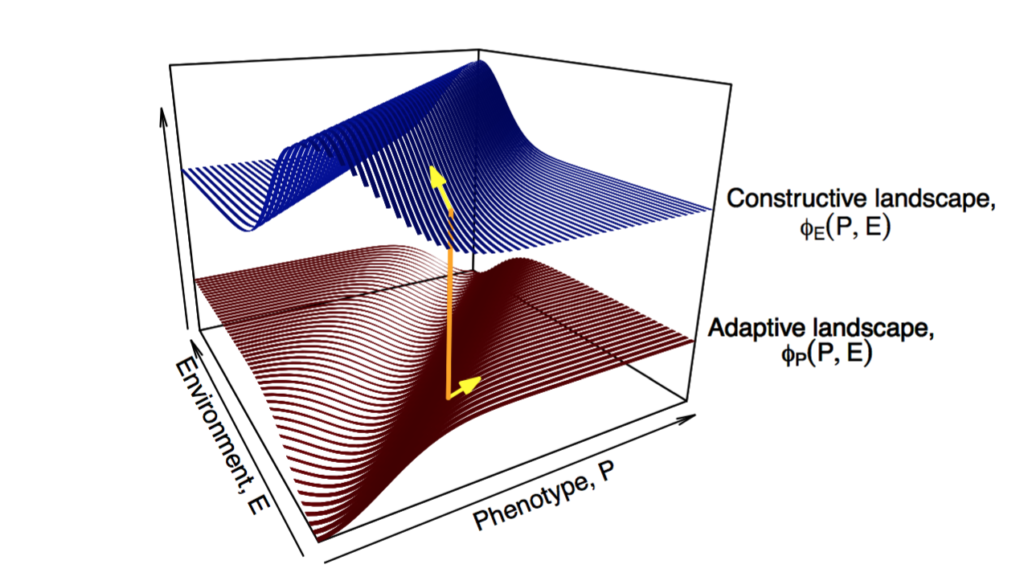
Organisms do not modify environments at random. Rather, they create environmental states, such as nests, burrows or benign conditions, that are adaptive for the constructor or its descendants. They can also destroy environments, or produce conditions that impact negatively on fitness. Studies show that environmental features constructed by organisms have different properties, and generate different patterns of selection, to aspects of environments that change independently of the organism (Clark et al. 2020). For instance, selection in response to environmental components regulated by organisms is consistently weaker and less variable than selection arising from autonomous aspects of environments. Organisms modify environments in distinctive, nonrandom ways, thereby imposing a systematic bias on natural selection.
The orderly nature of niche construction follows largely because it has evolved through earlier natural selection. However, this does not mean it can be disregarded. Organisms are influenced, but not determined, by their genes, and their activities are shaped by developmental information-gaining processes as well as natural selection. Organisms are not merely objects through which the causal explanatory power of natural selection flows; rather they are active agents that transduce and filter genetic inputs that derive from prior selection, as well as environmental inputs.
Fig 2 of Clark, Deffner, et al. 2020
(2) Niche construction generates ecological inheritance
Ecological inheritance refers to the inherited resources and conditions, and associated modified selection pressures, that ancestral organisms pass on to their descendants as a direct or indirect result of their niche construction. For instance, if a beaver builds a dam transforming its local ecology, the modified selection will remain in the beaver’s environment just so long as the dam, lake, and modified environment persist, which can be decades. Likewise, the changes that earthworms produce in the soil can last many generations and can increase the fitness of the descendants (see Blog: Evolution’s Engineers).
It is well-recognized that environments can exhibit constancy across generations, but this is not generally viewed as an inheritance. Yet these ecological legacies have been shown to affect evolutionary dynamics strongly, and to contribute to parent-offspring similarity (Odling-Smee et al. 2003; Badyaev & Uller 2009; Odling-Smee et al. 2013). The stable inheritance of traits results in part from parents constructing developmental environments for their offspring (Badyaev & Uller 2009).
In recent years many evolutionary biologists have sought to expand the concept of inheritance within evolutionary biology, and ecological inheritance is now commonly incorporated into these schemes (Danchin et al. 2011; Bonduriansky 2012).
(3) Learning can be evolutionarily significant
There is considerable interest among evolutionary biologists in the role that imprinting, song learning, habitat imprinting, cultural transmission and various other forms of learning, play in speciation, the evolution of adaptive specializations, adaptive radiations, the colonization of new habitats, brood parasitism and sexual selection in vertebrates (ten Cate 2000; Laland et al. 2019; Whitehead et al. 2019).
From the niche-construction perspective, acquired characters, such as learned behaviour, can be evolutionarily important. Social learning in particular, is likely to exert a widespread influence on animal evolution. For instance, different clans of killer whales feed on very different prey species, with individuals learning their dietary preferences from older group members. As a consequence, clans have evolved specific jaws and digestive systems adapted to cope with their learned diets. Killer whales may currently be evolving into multiple separate species because of their cultural differences (Foote et al. 2016).
The learning of one species can influence the evolution of another. Reed warblers, for instance, learn to recognize cuckoos as brood parasites by attending to the alarm calls of other birds, a knock-on consequence of which is that natural selection favors cuckoos with unusual plumage patterns (Thorogood & Davies 2012). This gene-culture coevolution is of particular relevance to human evolution (Laland et al. 2010).
Weed warblers and a cuckoo in the nest
(4) Byproducts are evolutionarily significant
The niche-construction perspective highlights the important roles that byproducts can play in ecosystems. Such roles are not intuitive. For instance, it is far more apparent that the beaver’s dam may drive coevolutionary episodes than beaver’s dung may, yet the latter is a very real possibility. Numerous examples have been documented of seemingly inconsequential and inadvertent acts by organisms whose aggregate activity generates important consequences. For example, consider Euchondrus snails whose consumption of endolithic lichens inadvertently generates tonnes of soil, thereby playing a vital role in desert ecosystems (Jones & Shachak 1990).
Typically, biologists assume that if a niche-constructing activity generates evolutionary feedback to the constructor, then it must be an adaptation. In fact, theory shows this need not be the case. Byproducts can induce selection on other traits in the same population and hitchhike to fixation on the back of this selection (Silver & Di Paolo 2006). Here, spatial structure (local dispersal and mating) gives rise to statistical associations between niche-constructing traits and genotypes favored in the constructed environments. There is selection of the niche-constructing trait, but not selection for it, and only the latter meets the definition of an adaptation (Williams 1966; Sober 1984). Nonetheless, in such hitchhiking cases, there remains evolutionarily consequential feedback to a niche-constructing population stemming from its constructing activities.
(5) Niche construction creates a second route to adaptation
Evolutionary theory has historically focused on how organisms are shaped by natural selection to become suited to their environments. The niche-construction perspective emphasizes that through niche construction, environments can be changed by organisms to suit themselves. For instance, Turner (2000) notes that, despite living on land for millions of years, earthworms have retained the physiology characteristic of the freshwater worms from which they evolved. The earthworms process the soil in ways that allow them to draw water into their bodies more effectively, constructing a simulated aquatic environment on land. The adaptive complementarity of earthworms and soils results to a large extent from the worms changing the soil through niche construction, rather than the worms evolving a typical terrestrial physiology through natural selection.
These findings have led to the claim that niche construction is more than just a product of evolution, or source of environmental change, but should be recognized as a causal evolutionary process through its guiding influence on selection.
(6) Niche construction opens up new ecological niches
Through their niche construction, organisms can open up new ecological niches, both for themselves and for other species. Experiments have shown that niche construction evolves rapidly, under a broad range of conditions (Callahan et al. 2014), often leading to the creation of new niches. For instance, San Roman & Wagner’s (2018) experimental evolution investigation in bacteria showed that huge biodiversity could emerge in a completely homogeneous environment through niche construction. They found that bacteria created new ecological niches when they excrete nutrient-rich waste products that could sustain other bacteria, and identified thousands of such niches that had been created in this manner. Rather than lineages simply diversifying to “fill” available niches, niches themselves may be diversifying (Erwin 2005). Recent theoretical work shows that the construction of new niches by organisms has long-term, macro-evolutionary effects, for instance, increasing the branching patterns of phylogenetic trees (Xue et al. 2020).
Clark AD et al. 2020. Niche construction affects the strength and variability of natural selection. The American Naturalist 195(1): 16-30. This meta-analysis of selection gradients provides clear evidence that the natural selection that arises from constructed environments differs from that from autonomous environments.
Odling-Smee FJ, Laland KN, Feldman MW 2003. Niche Construction: The Neglected Process in Evolution. Princeton: Princeton University Press An authoritative, rigorous and extensive introduction to niche construction theory.
San Roman M & Wagner A 2018. An enormous potential for niche construction through bacterial cross-feeding in a homogeneous environment. Shows experimentally that bacteria create new ecological niches when they excrete nutrient-rich waste products that sustain other bacteria
Mathematical evolutionary theory suggests that niche construction is of considerable ecological and evolutionary importance. It has been found to affect evolutionary equilibria, rates, and dynamics, drive coevolution, facilitate cooperation, and generate range expansion.
There is now an extensive body of formal theory that explores the evolution of niche construction and its ramifications for evolutionary biology and ecology. Mathematical analyses suggest that niche construction is of considerable ecological and evolutionary importance.
(vii) drive coevolutionary events, exacerbate and ameliorate competition, affect the likelihood of coexistence and produce macroevolutionary trends (e.g. Krakauer, Page & Erwin 2009).
(viii) strongly affect heritability, and challenges how it is commonly defined and measured (Uller & Helanterä 2020; see video on the right to learn more).
(ix) affect drift and gene flow, for instance, by creating suitable habitat that increases carrying capacities (Gurney & Lawton 1996), or through nonrandom dispersal which biases gene flow (Whitehead et al. 2019).
Creanza N, Feldman MW. 2014. Complexity in models of cultural niche construction with selection and homophily.Proceedings of the National Academy of Sciences USA. 111(3): 10830-7 Presents a model that includes selection and homophily as independent traits that influence the fitness and mate choice determined by another trait.
SilverM,Di PaoloEA. 2006. Spatial effects favour the evolution of niche construction.Theoretical population Biology.70:387-400A spatially explicit treatment of niche construction illustrates how niche-constructing traits can drive themselves to fixation in the absence of direct selection.
Researchers studying niche construction commonly embrace an alternative perspective in which niche construction is regarded as a fundamental evolutionary process in its own right. Evolution entails networks of causation and feedback in which previously selected organisms drive environmental changes, and organism-modified environments subsequently select for changes in organisms.
The extended evolutionary synthesis, or EES, is a new way to think about and understand evolutionary phenomena that differs from the conception that has dominated evolutionary thinking since the 1930s (i.e. the modern synthesis). It is a developing line of contemporary evolutionary thought that exists within the field of evolutionary biology.
The resulting network of processes provides a considerably more complex account of evolutionary mechanisms than traditionally recognized. The EES entails not only new research directions but also new ways to think about, and interpret, new and familiar problems in evolutionary biology.
Laland KN, Uller T, Feldman MW, et al. 2014 Does evolutionary theory need a rethink? Nature 514:161–164 Provides a very brief introduction to the motivation for an extended evolutionary synthesis.
Check out the Extended Evolutionary Synthesis resource website:
Why is niche construction contentious?
The fact that niche construction occurs is not at all contentious. Its study goes back to Darwin’s classic books on earthworms and corals, although the evolutionary consequences of niche construction have not always been fully appreciated. However, the study of niche construction has become contentious because theoretical and empirical findings from niche construction theory appear to challenge some orthodox accounts of evolution.
Researchers differ over to what extent niche construction requires changes in understanding of the evolutionary process, with many advocates of the niche-construction perspective aligning themselves with others seeking an extended evolutionary synthesis (Pigliucci & Muller 2010; Laland et al. 2014, 2015), a stance that other prominent evolutionary biologists reject (Wray et al. 2014).
The theoretical argument that lies at the heart of the claim that niche construction is an evolutionary process derives from the expectation that niche construction must be purposive and goal-directed, which follows from basics of thermodynamics. The great physicist, Erwin Schrödinger (1944) first pointed out that living organisms are far-from-equilibrium systems relative to their physical or abiotic surroundings. To preserve their out-of-equilibrium status, while creating order (structure, organization) in their bodies and their immediate surrounds, organisms must do work on their environment. Living organisms can only survive by constantly exchanging energy and matter with their …environments. Schrodinger explains why this work cannot be random; it must be directional. Random niche-constructing acts would not generate order in their bodies and immediate surrounds, and hence could not provide organisms with a basis for sustained life. It follows that organisms must change environments in systematic ways (Odling-Smee et al. 2003).
Over-and-above these thermodynamic considerations, niche construction generates environmental states that are coherent and integrated with the organism’s phenotype, and that are adaptive for the constructor or its descendants, largely because the organism’s capability to do so has evolved through earlier natural selection. Organisms in the same species possess broadly similar evolved capabilities for niche construction, so they are expected to modify environmental states in broadly similar ways, while the niche construction of offspring is expected to resemble that by their parents. For these reasons, niche construction theory expects niche construction to scale up across individuals in populations and over time to generate stable modification of environmental components, and as a result to generate consistent and sustained forms of natural selection. In this manner, niche construction imposes biases on selection.
This is probably the default assumption within evolutionary biology. There are two reasons why it is problematic. First, the niche construction of living organisms is only partly explained by earlier selection. Niche constructing capabilities can also arise in development.
For illustration, the fact that humans possess an evolved capacity to learn does not provide a complete and satisfactory explanation for why some humans populations happen to take up dairy farming, and hence why those particular populations evolved adult lactose tolerance through self-imposed selection on LCT alleles.
The second reason is the causal intertwining of evolutionary processes. Niche construction can only be reduced to past natural selection in a model of evolution in which the causes of Lewontin’s (1970) three conditions of ‘phenotypic variation’, ‘differential fitness’ and ‘inheritance’ are treated as quasi-independent: conversely, where these processes are causally intertwined, this interpretation does not follow (Uller & Helanterä 2017; Laland et al. 2019).
Niche construction is defined as ‘organism-driven environmental modification’. It thus includes any and all changes that an organism’s activities have upon the world. Niche construction, so characterized, is a universal feature of all living creatures. Even after death niche-constructing effects may persist: for instance, as a bodies decompose, they change the chemical composition of the earth around them, and sometimes leave hard parts, such as shells (Erwin 2008).
The generality of this formulation raises the question, ‘What is not niche construction?’ If almost everything an organism does is niche construction, is it still a useful concept? Yes – many biological concepts (growth, development, natural selection) apply universally, but that does not mean they are not useful. The correct question here is ‘When is it useful to emphasize that organisms engage in niche construction, and when is it not?
Some aspects of niche construction, for instance, the local fluctuations in ambient temperature caused by the movement of homothermic organisms, are frequently so trivial that they are best ignored. Incorporating these processes adds complexity but little explanatory power. Where organisms change their environments in very minor ways, or in ways that are immediately dissipated, negated, swamped or counteracted by other more powerful processes, such that they do not accrue across individuals or over time, it is better to ignore those aspects of niche construction.
What is not niche construction, then, are changes that organisms bring about in their worlds that are of no evolutionary or ecological consequence. This is an empirical issue. Whether discarded shells or hoof prints need to be treated as an instance of niche construction depends on whether the shells or indented soils provide resources for other organisms, and accrue in space and time, to affect selection on descendant populations.
Odling-Smee et al. (2003) defined niche construction as
‘the process whereby organisms, through their metabolism, their activities, and their choices, modify their own and/or each other’s niche’.
They defined an evolutionary niche, as ‘the sum of all the natural selection pressures to which the population is exposed’.
A feature of this characterization of niche construction is its breadth. The definition encompasses the construction of environmental modifications that have evolved as adaptations (such as birds’ nests) as well as environment-changing by-products (such as plant litter). It includes physical perturbations of the environment (e.g. digging a burrow) and relocations in space (e.g. migrations). It includes environment changing activities that have evolved through standard biological evolution as well as those that are learned and culturally transmitted (e.g. human agricultural practices).
Odling-Smee et al. coined this definition because they wanted to draw scientific attention to the diverse ways in which organisms modify environmental states, and the important ecological and evolutionary consequences that follow from these activities. What the above activities have in common is that they modify selection, as well as flows of energy and matter. A narrower definition risks neglecting important processes and feedbacks that fall outside it.
There is now extensive data showing that even organisms with modest individual effects on their environments can nonetheless strongly affect ecological and evolutionary events (Jones et al 1994, 1997; Odling-Smee et al. 2003; Sultan 2015). That is because a variety of processes scale up the impact of niche construction.
Organisms frequently repeat their niche-constructing activities across their lifespan. Organisms of the same population often modify their environments in similar ways, which means that even organisms with small per capita effects can have huge ecological effects where population densities are high. The effects of individual organisms can also accumulate in environments often over long periods of time, spanning multiple generations. In addition, some constructs or artifacts (such as termite mounds or beavers’ dams) are very durable or long-lasting, and can continue to have ecological consequences long after their constructors have died.
For all of these reasons, niche-constructing activities can act like a persistent unidirectional “pump” to eventually cause considerable change in environments.
No. The impacts that organisms have on environments are taken into account in countless evolutionary and ecological analyses. Ecologists have built large bodies of theory that explore the ecological consequences of many aspects of niche construction, for instance, models of resource depletion, succession, or competition for resources. Likewise, evolutionary biologists have developed models of frequency and density dependent selection, habitat selection, coevolution, sexual selection, maternal effects, and many other phenomena, in which the activities of organisms modify the action of selection. This literature is reviewed in considerable depth in Odling-Smee et al. (2003) and Sultan (2015), and is frequently discussed in articles and talks on niche construction.
Why then does Brodie (2005) write: ‘Natural selection is depicted [by niche construction theory] as resulting only from inanimate and abiotic features of the environment’. From where might this misunderstanding derive?
Possibly one source of confusion stems from a misreading of the above/adjacent figure, versions of which appear extensively in articles and Odling-Smee et al.’s (2003) book on niche construction. Here the horizontal arrows represent causal processes not effects in the world. Hence the absence of an arrow from organism to environment in figure 1a should be read as implying that niche construction is not recognized as an additional cause of evolutionary change, not that organisms are not recognized to alter environments.
This distinction between ‘process’ and ‘effect’ is important here. Evolutionary biology textbooks and articles frequently list a number of processes (selection, drift, mutation) that are causes of biological evolution, and invariably ‘niche construction’ is not found in such lists. The characterization of ‘niche construction’ as ‘the neglected process in evolution’ (Odling-Smee et al. 2003), implies only that it is not typically treated as process in evolution, and not that evolutionary biologists fail to recognize that niche construction happens.
Another potential source of misunderstanding may stem from the use of a pair of differential equations to illustrate the difference in perspective between niche construction theory and standard evolutionary theory. This heuristic dates back to Lewontin (1983), but has been reproduced widely (e.g. Odling-Smee et al. 2003).
Standard evolutionary theory is presented as dO/dt= f(O,E), dE/dt = g(E), but niche construction advocates argue that the following is more accurate: dO/dt= f(O,E), dE/dt = g(O,E).
It is possible to see how this characterization might be misleading, since on the surface it seems to deny any recognition by the standard account that environmental change can be caused by organisms. This was not Lewontin’s intention (personal communication to Kevin Laland).
Lewontin rejected the notion that the environment was ‘out there’ as an external context for evolutionary events that logically pre-exists the organism. The traditional accounts’ ‘externalism’ is well-recognized as a widespread metaphysical framework within biology by philosophers of science (e.g. Godfrey-Smith 1996).
Lewontin’s differential equations were never intended to apply to those aspects of the selective environment that constitute other organisms; for instance, sexual selection or coevolutionary theory. Nor did Lewontin’s equations deny ecological change caused by organisms. Rather, his equations were intended to challenge the standard separation of ecological and evolutionary processes and time-frames; the notion that one could study evolution without considering ecology, and vice-versa; that organismal effects on physical environments are too trivial or transient to exert any lasting impact; or even that a niche can be characterized without an organism.
Oxford evolutionary biologist Richard Dawkins (1982) introduced the term ‘extended phenotype’Biological adaptations expressed outside of the body of the organism. to represent the effects that genes can have outside of their bodies. Some, but not all, aspects of niche construction are also extended phenotypes. The latter is a narrower term because it is restricted to forms of environmental modification that are biological adaptations, such as birds’ nests or termites’ mounds.
In contrast, niche construction theory emphasizes that niche construction also incorporates evolutionary by-products. This broad characterization is vital, since in ecosystems the ecological and evolutionary consequences that flow from niche-constructing by-products are likely to be as substantial as those flowing from niche-constructing adaptations.
For instance, Post & Palkovacs (2009) describe how various patterns of guppy evolution are thought to have resulted from the different patterns of excretion observed in populations with different size distributions, while variant forms of root growth are observed in Populus trees and surrounding flora that have been differentially exposed to leaf litter with different tannin levels, since tannins impede nutrient release. They write: ‘eco-evolutionary feedbacks at the community and ecosystem level can emerge from both direct engineering and the by-products of living organisms (contrary to the arguments of Dawkins and Brodie).’
In addition, since extended phenotypes are restricted to biological adaptations, they automatically exclude aspects of niche construction that depend on acquired knowledge, for instance, human cultural processes. This is an important limitation for scholars of anthropogenic change, since very little human niche construction can accurately be characterized as a biological adaptation.
Most instances of human-induced habitat degradation – harvesting, deforestation, urban development, agricultural practices, livestock grazing, and so forth, – are products of cultural knowledge acquired and transmitted by humans, and expressed in their use of tools, technology and, engineering. In contrast to the extended phenotype, niche construction theory emphasizes how the acquired knowledge of organisms can trigger ecological cascades or drive evolutionary episodes.
Thirdly, extended phenotypes are properties of individuals, not of collections of individuals, yet some forms of niche construction are the results of actions by many populations, even many species. Both terrestrial and marine bioturbation influence the geochemistry and productivity of sediment, in turn influencing the evolutionary environment of a host of other species. The extended phenotype model is not a good description of the role of bioturbators, both because many effects are probably not adaptations, and because the sediment is a collective rather than individual product, comprised of the by-products of several species’ niche-constructing activity.
At the extreme, niche-constructing activities, such as sediment bioturbation or the accumulation of shell beds, can accumulate over geological time, modulating macroevolutionary patterns and diversity (Erwin 2008). This temporal dimension to niche construction, in shaping the pattern and intensity of selection experienced by distant descendants, is entirely missed by the theory of the extended phenotype. However, numerous theoretical models have shown that it strongly affects evolutionary dynamics
Richard Dawkins (2004) criticized niche construction theory for ‘confusing’ byproducts and adaptations:
‘The problem I have with niche construction is that it confuses two very different impacts that organisms might have on their environments. As Sterelny (2001, p. 333) put it, ‘Some of these impacts are mere effects; they are byproducts of the organisms’s way of life. But sometimes we should see the impact of organism on environment as the organism engineering its own environment: the environment is altered in ways that are adaptive for the engineering organism. Niche construction is a suitable name only for the second of these two (and it is a special case of the extended phenotype). There is a temptation, which I regard as little short of pernicious, to invoke it for the first (byproducts) as well.’
In fact, niche construction theory does not ‘confuse’ adaptations and byproducts. Scientific articles regularly distinguish between niche-constructing adaptations and niche-constructing byproducts.
The difference in perspective stems from the fact that Dawkins’ focus is on the evolution of niche-constructing traits, where his distinction between adaptations and byproducts is most relevant. However, niche construction theory has a broader focus that also encompasses the ecological and evolutionary consequences of niche construction for other species, where this distinction is only sometimes relevant.
Dawkins’ extended-phenotype stance, in restricting evolutionary feedback to the alleles underlying a niche-constructing trait only, may arguably be adequate as a conceptual simplification with which to understand the evolution of a specific niche-constructing trait (although even here the extended phenotype position may overlook important processes). However, Dawkins’ extended phenotype stance ignores additional forms of feedback that flow to the constructor, and to other populations in its local environment, from its niche-constructing activity, indirectly via the environment.
In practice, it is frequently (almost always) not known whether an instance of niche construction is an adaptation or a byproduct. Demonstrating that something is an adaptation is a very challenging thing to do. For this reason, many researchers find a neutral umbrella term like ‘niche construction’ useful in practice.
For the evolution ofniche construction the appropriate focus is on adaptations, and on the direct and indirect fitness benefits accrued to niche constructors as a result of modifying their environment. However, for the ecological or evolutionary consequences ofniche construction (for instance, the evolution of other traits, or of other populations, or on ecological flows and resource distributions, or eco-evolutionary feedbacks, or on the structure and characteristics of ecosystems), it is vital that analysis is not restricted to adaptations, but includes the many important consequences that flow from time-lagged effects, by-products, acquired characters, and collective activity.
‘Niche construction is not just an important addition to current evolutionary theory; it requires a reformulation of evolutionary theory.’ (Odling-Smee et al. 2003)
‘The changes to the evolutionary process brought about by niche construction… are sufficiently important and occur sufficiently frequently to warrant an overhaul in evolutionary thinking.’ (Day et al. 2003)
Claims, such as the above, made about niche construction by its advocates, have been viewed as extravagant overstatements, hyperbole or ‘hard sell’ by its critics. Conversely, advocates view such statements as justified, on the grounds that the refinement in evolutionary thinking that they advocate is genuinely radical.
For instance, the inclusion of a general second inheritance system (ecological inheritance), the explicit recognition of the role of acquired characters in directing evolution, and the emphasis on reciprocal causation, are three ways in which niche construction theory differs from standard accounts quite distinctly.
Niche construction theory describes the evolutionary process in a fundamentally different way. If an advocate draws the conclusion that their science is better served by recognizing niche construction as a distinct evolutionary process then to portray it as a radical departure is not exaggeration.
The claims that niche construction is an evolutionary process, a source of adaptation, and requires an extended evolutionary synthesis, have all excited controversy. These issues are discussed by Laland & Sterelny (2006), and by Scott-Phillips et al. (2014). The latter article is a collaboration between some critics of the niche-construction perspective and one of its advocates that attempts to pinpoint their differences. The authors conclude that their disagreements reflect a wider dispute within evolutionary theory over whether the neo-Darwinian synthesis is in need of reformulation, as well as different usages of some key terms (e.g., evolutionary process). Finally, Uller & Helanterä (2017) discuss the different conceptions of biological causation that separate niche construction advocates and critics.
Laland KN, Odling-Smee FJ, Feldman MW. 2019. Understanding niche construction as an evolutionary process. In Uller T & Laland KN, eds. Evolutionary causation. Biological and Philosophical Reflections. Cambridge, MA: MIT Press. Discusses why niche construction theory is well-received in some academic fields and contentious in others.
Laland KN, Sterelny K. 2006. Seven reasons (not) to neglect niche construction.Evolution. 60: 1751–1762. Discusses seven reasons given for not recognizing niche construction as evolutionarily important, taking issue with these.
Odling-Smee FJ, Laland KN & Feldman MW 2003. Niche Construction: The Neglected Process in Evolution. Princeton University Press. An authoritative, rigorous and extensive technical introduction to niche construction theory. This monograph discusses some points of controversy.
Scott-Phillips TC, Laland KN, Shuker DM, Dickins TE & West SA (2014) The niche construction perspective: a critical appraisal. Evolution 68(5): 1231-43. A useful exchange between some critics of the niche-construction perspective and one of its advocates that is successful in pinpointing some of their differences.
Uller T, H Helanterä. 2019. Niche construction and conceptual change in evolutionary biology. British Journal for the Philosophy of Science. 70:2. 351-375. Explains how receptive or non-receptive attitudes to niche construction theory follow from assumptions about whether Darwinian processes are causally independent.
Walsh DM. 2015. Organism, Agency and Evolution. Cambridge: Cambridge University Press. Discusses how and why contemporary evolutionary biology neglects the agency of organisms.
Multiple routes to adaptation
Organisms can respond plastically to novel conditions to generate functional variation. The specific adaptive phenotypes generated need not be the direct targets of past selection. Rather, they are often the expression of the more general ability of developmental processes to accommodate novel inputs adaptively, enabling functionally integrated responses to a broad range of conditions (West-Eberhard 2003; Kirschner & Gerhart 2007).
Moreover, through niche construction, environments can be changed by organisms to suit themselves. Consider the example of the earthworms which, despite living on land for millions of years, have retained the characteristic physiology of the freshwater animals from which they evolved (Turner 2000). For instance, earthworms produce large quantities of dilute urine, a trait that on land leaves them highly vulnerable to desiccation. Earthworms can only survive because they process the soil in ways that allow them to draw water into their bodies more effectively. They construct a simulated aquatic environment on land.
Fig 7.1 from Turner 2000
The key point is that the adaptive complementarity of earthworms and soils results to a large extent from the worms changing the soil through niche construction, rather than natural selection changing the worms to a typical terrestrial physiology.
Traditionally, the latter route has not been recognized as an alternative to adaptation – after all, the earthworm’s niche-constructing traits are adaptations too. However, the conventional stance assumes a particular model of biological causation, which advocates of the niche-construction perspective reject (Laland et al. 2011).
From the niche-construction perspective, the current selective environment was itself brought about through earlier niche construction, which caused the bout of selection. The attribution of all causal significance to selection fails to capture the true reciprocal nature of causation in this system.
Central to these debates are different assumptions concerning the independence or interdependence of the causes of phenotypic variation, differential fitness and inheritance, which are Lewontin’s (1970) three conditions for evolution by natural selection (Walsh 2015; Uller & Helanterä 2019). Traditionally, evolutionary biologists have assumed these processes are quasi-independent, but in practice they are often causally intertwined.
Contents
Causal Interdependencies
An orthodox stance would not recognize earthworm niche construction as an evolutionary cause but rather treat it as the outcome of earlier natural selection that favored earthworm soil-processing capabilities.
Implicit in this reasoning is the separation of the processes that generate variation (e.g., mutations in soil-processing activity) and the causes of fitness differences (e.g., the alternative environmental conditions that favored those mutations). This allows the explanation for the adaptive fit between earthworms and their soil environment to begin with that change in ancestral environmental conditions that generated selection for earthworm soil processing.
For instance, in the figure on the right sidebar, (1) a random mutation, (2) changes the earthworms’ soil processing, and (3) is selected because of beneficial effects on fitness.
In reality, because ancestral earthworm activity is itself the cause of the soil environment that favors mutations in soil processing, the causes of variation and fitness differences are not independent (bottom figure). The explanation for the adaptive fit between earthworms and their soil environment cannot begin with those ancestral external environmental conditions that elicited selection for earthworm soil processing, since those conditions were themselves products of earlier earthworm niche construction.
Worse, the processes underlying inheritance are also not independent of the causes of variation and fitness. The genetically specified propensity for soil processing that contemporary earthworms inherit only functions effectively as the source of earthworm adaptation to a soil environment because contemporary earthworms also inherit the ecological legacy of a modified soil environment that is the product of ancestral earthworm niche construction.
Traditionally, this causal interdependence has been neglected or downplayed. Recognizing niche construction as an evolutionary process is how niche construction theory builds causal interdependence into evolutionary explanation.
Harvard evolutionary biologist and Modern Synthesis architect Ernst Mayr (1961) wrote about biological causation, distinguishing between proximate and ultimate causes. Mayr equated proximate causation with immediate factors (e.g. physiology) and ultimate causation with evolutionary explanations (e.g. natural selection). He argued that proximate and ultimate causes addressed different questions and were not alternatives, which remains a valid and non-contentious point.
Mayr’s account of causation remains widely accepted today, with both positive and negative ramifications. One negative consequence was that Mayr insisted on a strict separation of proximate and ultimate causes:
“The clarification of the biochemical mechanism by which the genetic program is translated into the phenotype tells us absolutely nothing about the steps by which natural selection has built up the particular genetic program” (Mayr 1980).
This position was based upon a specific view of development:
“All of the directions, controls and constraints of the developmental machinery are laid down in the blueprint of the DNA genotype as instructions or potentialities” (Mayr 1984).
Mayr and his followers argued that developmental (i.e. ontogenetic) processes were irrelevant to the study of evolution. Niche construction is an ontogenetic process, and advocates of the niche-construction perspective, alongside others, have taken issue with Mayr’s treatment of causation (Laland et al. 2011).
For Mayr, developmental processes could not be regarded as independent causes of evolutionary events, since he believed their properties (which include how organisms construct their niches) were themselves fully explained by prior natural selection. From Mayr’s standpoint, if developmental processes direct evolutionary events, this is only the proximate manifestation of the ultimate cause of natural selection; aspects of development not shaped by selection were assumed to play no evolutionary role.
This stance has made it more difficult for evolutionary biologists to recognize niche construction as an evolutionary process. Niche construction is perceived to have no independent evolutionary significance because, to the extent that it is evolutionarily consequential, it is regarded as fully explained by a preceding cause, natural selection (e.g. Dawkins 2004). Niche-construction effects are treated as ‘extended phenotypes’, whose sole evolutionary role is to affect the probability that gene variants underlying niche construction are passed on to the next generation. Similar reasoning underlies the treatment of niche construction as an indirect genetic effect, with, for example, a mother’s genes affecting her offspring’s phenotype by modifying its environment (Wolf et al. 1998).
There are two major problems with this line of reasoning:
First, as described above, the processes underlying variation, fitness and inheritance are causally intertwined. All organisms have always engaged in niche construction, going right back to the beginning of life. Hence there is no reason to assume that natural selection must be the first cause of any evolutionary episode. Causation in biological systems is reciprocal rather than linear, with natural selection and niche construction codirecting each other.
Second, not all evolutionarily consequential aspects of niche construction (nor all aspects of development, in general) are under genetic control. Niche-construction theorists, like many developmental biologists, regard organisms (and their environmental modification) as under-determined by genes. The changes that organisms bring about do not flow only from their adaptations, but also derive from plasticity, byproducts, and acquired characters.
Laland KN, Odling-Smee FJ, Gilbert SF. 2008.EvoDevo and niche construction: building bridges.Journal ofExperimentalZoology Part B. 310:549–566. Points to common ground between niche construction and evo devo. Argues that the same conceptual barriers have hindered both.
Laland KN, Odling-Smee FJ, Feldman MW. 2019. Understanding niche construction as an evolutionary process. In Uller T & Laland KN, eds. Evolutionary causation. Biological and Philosophical Reflections. MIT Press. Discusses why niche construction theory is well-received in some academic fields and contentious in others.
The agency of organisms
Waddington and Lewontin both objected to any portrayal of organisms as passive victims of selection. They viewed organisms as active agents, both in their own development and in their evolution, with their activities a major determinant of fitness differences and hence of natural selection.
Agency is the intrinsic capacity of individual living organisms to act on, and in, their world, and thereby to modify their experience of it, including in ways that are neither predetermined, nor random (Laland et al. 2019). It is an essential and inescapable aspect of nature. Living organisms are not just passively pushed around by external forces, but rather they act on their world according to intrinsically generated but historically informed capabilities. Organisms are self-building, self-regulating, highly integrated, functioning, and (crucially) “purposive” wholes, which through wholly natural processes exert a distinctive influence and a degree of control over their own activities, outputs, and local environments. Indeed, organisms must have these properties in order to be alive (Schrödinger 1944).
Use of the term ‘agency’ in niche construction theory does not imply conscious, sentient, or deliberate action, nor vitalism (or any mystical power that imbues living tissue), nor a rejection of mechanistic explanation, nor the belief that living organisms possess any desire to evolve, or achieve some final state. Human niche construction sometimes is conscious, intentional, and deliberate, and may set out to achieve longer-term goals, but these are neither necessary nor defining characteristics of niche construction. Even for humans, a great deal of our niche construction is non-intentional. The claim that organisms are “purposive” means nothing more than that organisms exhibit goal-directed activities, such as foraging, courtship, or phototaxis, which are entirely natural tendencies with short-term local objectives, and that have themselves evolved.
In acknowledging the legacy of past natural selection in shaping how contemporary organisms manifest their agency in the world, niche construction theory rejects the suggestion that the actions of organisms are fully explained by prior selection. Organisms are influenced, but seldom if ever fully determined, by their genes, and their activities are shaped by developmental information-gaining processes as well as selection. They are not merely objects through which the causal explanatory power of natural selection flows; organisms are active agents that transduce and filter genetic and environmental inputs.
Laland KN, Odling-Smee FJ, Feldman MW. 2019. Understanding niche construction as an evolutionary process. In Uller T & Laland KN, eds. Evolutionary causation. Biological and Philosophical Reflections. MIT Press. Discusses why niche construction theory is well-received in some academic fields and contentious in others. The issue partly depends on disciplinary attitudes to agency.
WalshDM. 2015. Organism, Agency and Evolution. Cambridge: Cambridge University Press. Discusses how and why contemporary evolutionary biology neglects the agency of organisms.
Evolutionary theory is often viewed as a fusion of Darwinian and Mendelian insights. Notwithstanding the immense progress that these achievements engendered, Richard Lewontin (1983) argues that the legacies of Darwin and Mendel include misleading metaphors that:
are responsible for certain difficulties in biology, difficulties that prevent us from some kinds of further progress and which keep us locked into a rigid framework of thought about the development and evolution of organisms.
Mendel’s view of organisms as the manifestation of autonomous internal ‘factors’ (i.e. genes) with their own laws, germinated into a post-Synthesis metaphor in which ontogeny ‘is seen as an unfolding of a form, already latent in the genes, requiring only an original triggering at fertilization and an environment adequate to allow ‘normal’ development to continue’. We now know that developmental processes are hugely environmentally contingent.
Darwin’s view of organisms as passive objects moulded by the external force of natural selection encouraged a conception of evolution in which ‘the environment “poses the problem”; the organisms posit “solutions”, of which the best is finally “chosen”. The metaphor of selection, inspired by the efficacy of artificial selection, continues to encourage a view of organisms as passive objects upon which external forces act.
Lewontin argues forcefully that these metaphors are misleading. Similar points have been made by Oyama (1985), Griffiths and Gray (2001, 2004), Sultan (2015) and many others.
Lewontin (1983) also took issue with the adaptive landscape metaphor, which is widely used to visualise the fitness of organisms as a function of genetic or phenotypic state. Evolving organisms are general construed as climbing the fitness landscape to a local peak through the action of natural selection. In practice, landscapes are not static. Lewontin pointed out that through their niche construction the organisms themselves alter the landscape, so that moving across a landscape is like walking on a trampoline.
Recently, Tanaka et al (2020) proposed a dual-landscape model of adaptation with natural selection and niche construction, and used it to analyse the dynamics of three cases of microbial evolution.Tanaka et al’s (2020) dual landscape model of evolution with natural selection and niche construction. Reproduced from Figure 2.
Lewontin RC. 1983. Gene, organism, and environment. In: Bendall DS, ed. pp. 273-285. Cambridge: Cambridge University Press. This accessible essay introduces the niche construction perspective and critiques conventional evolutionary approaches to adaptation.
Levins R, Lewontin RC. 1985. The Dialectical Biologist. Cambridge: Harvard University Press. Discusses how metaphors concerning selection and development can be misleading.
Niche construction in ecology
What is a niche?
A key feature of modern niche concepts is that the ecological niche is defined relative to the organism at its centre. This is because organisms determine which environmental factors are significant components of their world, a perspective very much in line with niche construction theory.
Organisms actively modify their niches, and those of other organisms, through their activities and choices. However, niches have typically been construed as static rather than evolving. In order to devise a more dynamic niche concept, Odling-Smee et al. (2003) characterize an evolutionary niche as ‘the sum of all the natural selection pressures to which the population is exposed’, an approach well-received by ecologists (Chase & Leibold 2003). The evolutionary niche can be viewed as a dynamic (i.e. evolving) version of the more familiar ecological niche when the latter is characterized as an n-dimensional hypervolume of those conditions and resources relevant to the population (Hutchinson 1957).
How do organisms modify niches?
Organisms change their niches, by physically changing environments, or by relocating in space, thereby exposing themselves to new conditions. They may create novel niches by inceptive niche construction, by innovating. For example, the evolution of improved paper technology in Polistinae wasps apparently had massive effects on the geographic distribution, colony size and social complexity of these wasps (Hansell 1993).
Nest building has shaped the evolution and distribution of wasps.
Organisms can also buffer out some of the changes in their environments that are caused by other agents by counteractive niche construction. For instance, wasps regulate temperature in their nests by bringing droplets of water to cool the nest, or by stretching and contracting their abdomens in unison to warm the nest.
Through niche construction, organisms can influence and control some flows of energy and materials among trophically interconnected organisms, even without being part of those flows themselves. The modification of natural selection by niche construction may be direct, as occurs when animals construct artifacts, such as nests or dams. Conversely, organisms may only modify natural selection indirectly, or gradually, for instance through the slow accumulation in their environments of the by-products of their metabolisms, and detritus producing activities – which generates an ecological inheritance.
How does niche construction relate to ecosystem engineering?
By building dams, beavers modify river ecology for decades, dramatically changing the environments of numerous organisms.
Jones et al. drew attention to the comparative lack of ecological research dedicated to studying organisms that modulate the availability of resources and habitats in ecosystems. For instance, beavers’ dams have dramatic effects on the flows of energy and matter through river environments. Kelp are another example: their growth creates forests that provide habitat for countless other organisms.
Kelp forests create homes for countless marine creatures.
Many species do influence energy flows, mass flows, and trophic patterns in ecosystems, by generating ‘engineering’ control webs based on mosaics of connectivity among species . Jones et al. have shown that engineering webs make significant contributions to the regulation of energy and matter flows in ecosystems, alongside webs of trophic interactions.
How are ecologists using niche construction theory?
Recent years have witnessed considerable efforts to operationalize the niche construction concept within ecology, and devise means for its investigation. These methods provide tools to explore how niche construction triggers ecological and evolutionary feedbacks, to detect the ecological signatures that it leaves, and to explore its effects on biodiversity.
Post & Palkovacs (2009) illustrate how niche construction can generate eco-evolutionary feedbacks, and how these can be investigated.
Table 1 from Post and Palkovacs 2009
Odling-Smee et al. (2013) describe some of the ecological and evolutionary impacts on ecosystems of niche construction.
Matthews et al. (2014) propose an operational framework to evaluate comparative and experimental evidence of the evolutionary consequences of niche construction, and suggest how such research can improve our understanding of ecological and evolutionary dynamics in ecosystems .
Brathen & Raivolainen (2015) show that species sharing a single trait or species belonging to a growth form can act as collective niche constructors, and be important predictors of species diversity in ecological communities. In tundra plant communities, forbs and grasses were the least abundant growth forms, yet they were the strongest positive predictors of species diversity.
Ware et al. (2019) propose a conceptual model that shows how feedbacks across levels of organization link theory associated with eco-evolutionary dynamics, niche construction and the geographic mosaic theory of evolution.
Fig 1 from Ware et al. 2019
Coexistence, range expansion and adaptive radiation
Theoretical studies show that niche construction has important effects on species coexistence, range expansion and adaptive radiation.
Kylafis & Loreau (2011) introduce the concept of ‘ecological niche construction’, the process whereby an organism improves its environment to enhance its growth and persistence, which they argue is an important missing element of niche theory. In a model of two consumers that compete for one limiting resource and one predator, they show how niche construction modifies the traditional niche-deteriorating impacts of its agent or of competing species, and hence the potential for species coexistence. Thus niche construction can promote mutualism and niche partitioning, although under different conditions it can strengthen interspecific competition. The authors also show how niche construction strongly affects the realized niche of a species.
Kylafis & Loreau (2008) demonstrate that niche construction can generate local spatial effects that can allow a population to promote its own range expansion. In this manner, niche construction potentially plays a role in adaptive radiation. Such effects can link eco-evolutionary feedbacks to the process of adaptive radiation. Rather than lineages simply diversifying to “fill” available niches, niches themselves may be diversifying (Erwin 2005), a process subsequently termed “self-propagating adaptive radiation.”
Krakauer, Page, & Erwin (2009) show that niche construction can drive coevolutionary events, exacerbate and ameliorate competition, affect the likelihood of coexistence and produce macroevolutionary trends.
Buser CC, Newcomb RD, Gaskett, AC & Goddard MR. 2014. Niche construction initiates the evolution of mutualistic interactions. Ecology letters. 17(10): 1257-1264. Demonstrates experimentally how through niche construction (modification of fruit) the yeast Saccharomyces cerevisiae attracts Drosophila, facilitating its propagation.
Erwin DH . 2005. Seeds of diversity.Science. 308:1752–1753. Presents the hypothesis that organisms construct new niches, leading to diversification.
Hamblin SR, White PA, Tanaka MM. 2014. Viral niche construction alters hosts and ecosystems at multiple scales. Trends in Ecology & Evolution.29(11): 594-9. Viruses modify host environments, and these modifications drive evolutionary feedback between the virus and its environment across multiple scales from cells to ecosystems.
Hansell MH. 1993. The ecological impact of animal nests and burrows.Functional Ecology. 7: 5–12. Provides extensive empirical examples of the ecological impact of niche construction through nest and burrow construction.
Jones CG, Lawton JH, Shachak M. 1994. Organisms as ecosystem engineers.Oikos. 69: 373-386. The most authoritative introduction to the concept of ecosystem engineering.
Krakauer DC, Page KM, Erwin DH. 2009. Diversity, dilemmas, and monopolies of niche construction.American Naturalist. 173: 26–40. Demonstrates a fundamental dilemma of niche construction, whereby the construction of a shared resource leads to a tragedy of the commons, with competition tending to eliminate niche construction strategies.
Kylafis G, Loreau M. 2008 Ecological and evolutionary consequences of niche construction for its agent.Ecology Letters. 11: 1072-1081. This theoretical analysis shows that this niche construction allows the persistence of the plants under infertile soil conditions that would otherwise lead to their extinction.
Kylafis G, Loreau M. 2011 Niche construction in the light of niche theory.Ecology Letters. 14: 82–90. Introduces the concept of ‘ecological niche construction’, the process whereby an organism improves its environment to enhance its growth and persistence.
Matthews B, De Meester L, Jones CG, Iberlings BW, Bouma TJ, Nuutinen V, van der Koppel J & Odling-Smee J. 2014. Under niche construction: an operational bridge between ecology, evolution and ecosystem science. Ecological Monographs. 84.2: 245–263. Written for professional ecologists with a view to operationalizing niche construction, this article illustrates how niche construction can be investigated.
Odling-Smee FJ, Erwin D, Palkovacs E, Feldman M, Laland KN. 2013 Niche construction theory: a practical guide for ecologists.Quarterly Review Biology. 88, 3-28. This article provides a practical guide to how niche construction theory can be deployed by ecologists and evolutionary biologists to explore eco-evolutionary dynamics.
Why has niche construction theory been so influential in the human sciences? One reason is that it is self-apparent that humans possess an unusually potent capability to regulate, construct and destroy environments, and that this is generating some pressing current problems (e.g. climate change, …deforestation, urbanization). A second reason is niche construction theory’s recognition of human agency (and of human activities guiding selection rather than merely resulting from it), which is attractive to human scientists (Odling-Smee et al. 2003; Kendal et al. 2011; O’Brien & Laland 2012), with the important caveat that this emphasis on agency does not imply human niche construction is necessarily conscious or deliberate. A third reason is that niche construction theory emphasizes how acquired characters play an evolutionary role, and this is particularly relevant to human evolution, where our species appears to have engaged in extensive environmental modification through cultural practices (Laland et al. 2010). Niche construction theory provides an evolutionary framework capable of accommodating human cultural behavior in a way that doesn’t diminish its complexity and uniqueness, yet also shows how these behaviors are an extension of general evolutionary processes that pertain to non-human organisms.
Such cultural activities are typically not themselves biological adaptations (rather, they are the adaptive product of those much more general adaptations, such as the ability to learn, particularly from others, to teach, to use language, and so forth, that underlie human culture) and hence, cannot accurately be described as extended phenotypes.
Mathematical models reveal that niche construction due to human cultural processes can be as potent as gene-based niche construction, and establish that cultural niche construction can modify selection on human genes and drive evolutionary events (Laland et al. 2001; Creanza et al. 2012; Creanza & Feldman 2014, 2016). This interaction, known as ‘gene-culture co-evolution’, is also found in some animals, such as killer whales, orangutans and reed warblers (Whitehead et al. 2019). However, gene-culture coevolution reaches its zenith in humans. Cultural change will typically occur faster than genetic adaptation, and this has allowed cultural niche construction to play a prominent role in human evolution (Creanza et al. 2017).
Cultural traditions may be driving speciation in killer whales
There is now little doubt that human cultural niche construction has co-directed human evolution (Laland et al. 2010; Creanza et al. 2017). Humans have modified selection, for instance, by dispersing into new environments with different climatic regimes, devising agricultural practices or domesticating livestock.
The best-researched example of gene-culture co-evolution is the finding that dairy farming created the selection pressure that led to the spread of alleles for adult lactase persistence (Feldman & Cavalli-Sforza 1989; Gerbault et al. 2011). However, analyses of the human genome have identified many hundreds of genes subject to recent selection, many in response to human cultural activities (Laland et al. 2010). The evolution of lactose persistence may be representative of a very general pattern of gene-culture coevolution over the last 20,000 years.
One study reported 27 separate genes, known to have been subject to recent selection, for which the inferred cultural selection pressure is a change in diet associated with the advent of agriculture (Laland et al. 2010). In addition to the breakdown of dairy products, the list also includes genes expressed in the metabolism of carbohydrates, starch, proteins, lipids, phosphates, plant secondary compounds and alcohol, as well as jaw muscle fibres and tooth-enamel thickness.
Table 2 from Laland et al. 2010
In some West African populations, the cultural niche construction practice of cultivating yams has inadvertently generated selection for the haemoglobin S allele that underlies sickle cell anaemia. Slash and burn agriculture creates conditions that lead to increased standing water when it rains, and these puddles are perfect breeding grounds for malaria carrying mosquitos. The enhanced incidence of malaria, in turn, generates selection for gene variants that confer resistance to malaria, one of which is Hb S.
Cultural niche construction can also feed back to influence the cultural evolution of a second trait, for instance allowing for the coevolution of marriage customs and sex ratio biases (Creanza et al. 2012). Here, the spread of a cultural trait creates a cultural niche in which another cultural trait flourishes. A good example is provided by the demographic transition. The reduction in birth rate during the demographic transition is often viewed as a paradox because, from a traditional evolutionary perspective, it is difficult to envisage why individuals should prefer to have fewer children. However, cultural niche construction models show that if a cultural norm favoring greater education spreads, a preference for smaller family size follows, allowing the fertility rate to drop (Ihara & Feldman 2004).
The primary cause of cultural complexity is another conundrum that niche construction theory has helped to resolve. The issue had proven contentious, with some researchers suggesting that large populations support diverse cultural knowledge and others arguing that environmental factors are more important. The paradox is resolved when niche construction is taken into account (Collard et al. 2012; Fogarty & Creanza 2017). The effects of a changeable environment, to which food gatherers are subject, can be attenuated by the niche-constructing activities of food-producers, such as agriculturalists. As a result, the effect of the environment on cultural complexity is stronger in food-gathering populations and weaker in food-producing populations, whilst the effect of population size is stronger in producers compared to gatherers.
The niche-construction perspective has been productive in many other studies of human social behavior (Kendal et al. 2011; Creanza et al. 2017), including some that describe some dramatically large-scale consequences of culturally driven change. These include the extinction of megafauna after the arrival of humans and consequent shaping of global species distributions (Boivin et al. 2016), and the striking changes of landscapes in Bali (Lansing et al. 2009; Lansing & Fox 2011) and Polynesia (Quintus & Cochrane 2018) with complex cascades of ecological and social consequences following the cultivation of crops.
The human transition from hunting and gathering to food production economies provides another illustration of the explanatory power of niche construction theory (Smith 2007a, 2007b, 2012, 2016). Archaeological and paleo-environmental records from eastern North America, Amazonia, the Near East, and, increasingly, China,contradict the assumption of traditional explanatory frameworks that environments change and species adapt (Smith 2011; Zeder 2017; Ren et al. 2016; Lombardo et al. 2020). Conversely, explanations based on niche construction theory are well-supported in these regions. Archaeologists have concluded that extensive data concerning the domestication of plants and animals, and the development of agricultural societies, supports predictions derived from niche construction theory (Zeder & Smith 2009; Zeder2017, 2018; Piperno 2017). Initial domestication of plants and animals, in turn, offer proponents of niche construction theory and the EES an opportunity to evaluate core assumptions about niche construction, ecological inheritance, plasticity first evolution, and reciprocal causality, in a major transition with profound impact on both human and non-human organisms (Zeder 2018).
Psychologists and linguists are also using niche construction theory. Researchers have stressed that the human mind is a symbol-generating and artefact-devising system, as a result of which children develop in a rich human-constructed world teeming with diverse symbols and artefacts. The constructed cognitive niche both shapes, and is shaped by, the child’s learning and development (Flynn et al. 2013). Peterson et al (2018) argue that physical signs and symbols left in the environment have played an important role in human evolution.
Niche construction is also now central to several accounts of how language evolved. For instance, Bickerton (2009) describes how our ancestors constructed scavenging niches that required them to communicate in order to recruit sufficient individuals to drive off predators away from megafauna corpses. He maintains that our use of language, in turn, created a new niche in which sophisticated cognition was beneficial. Tomlinson (2015, 2018) makes a related argument, and shows how it explains the origins of human music.
Anton SC, Potts R, Aiello LC. 2014. Evolution of early Homo: An integrated biological perspective.Science. 345(6192): 1236828. A recent overview of current thinking in the field of human evolution. Niche construction is thought to play a central role.
Flynn EG, Laland KN, Kendal RK, Kendal JR. 2013 Developmental niche construction. Developmental Science. 16(2): 296-313. Draws attention to parallels between niche construction theory and some fields of developmental science, and illustrates now the niche-construction perspective can be fruitfully applied within developmental psychology.
Gerbault P, Liebert A, Itan Y, Powell A, Currat M, Burger J, Swallow DS, Thomas MG 2011. Evolution of lactase persistence: an example of human niche construction. Phil. Trans. R. Soc. B. 366: 863–877. The domestication of cattle and consumption of dairy products is a compelling human example of niche construction, which has selected for alleles for adult lactose absorption.
Kendal JR, Tehrani JJ & Odling-Smee FJ. 2011. Human niche construction.Philosophical Transactions of the Royal Society B. 366: 1566. A series of articles on human niche construction.
O’Brien M, Laland KN. 2012. Genes, culture and agriculture: an example of human niche construction.Current Anthropology. 53: 434-470. Draws on niche-construction theory and gene-culture co-evolutionary theory to propose a broad theoretical framework with which archaeologists and anthropologists can explore human co-evolutionary dynamics.
Niche construction arises in development: it occurs through the activities of individual organisms during their lifetime (ontogeny) as they modify environmental states, and has ecological and evolutionary consequences when those individual-based local effects scale up across populations, and over time. By modifying their local environment, organisms can affect their own future development, whilst the construction of nests and other ‘nursery’ environments can shape and regulate the conditions experienced by developing offspring. For these reasons, niche construction theory is explicit about the need to treat niche construction as both a developmental process and a cause of evolution (Odling-Smee et al. 2003).
There exists a complementarity between the fields of niche construction and ecological developmental biology (Laland et al. 2008). Niche construction emphasizes the ability of the organism to modify the environment to form a supportive niche, whilst ecological developmental biology emphasizes the ability of environmental agents (e.g. temperature, density, symbionts) to modify the developing organism.
Animal and plant development provide some of the most direct examples of niche construction (Gilbert 2020). Here, two modes of niche construction can occur, often simultaneously. First, the developing organism can modify its environment to construct niches for its own development. Second, the organism can become the niche for other organisms that will later alter it. Both of these modes involve developmental plasticity–the ability of an organism to change its phenotype in response to the presence of environmental agents.
Developing organisms modify their own environments to construct niches
The formation of galls by gall wasps and the formation of the placenta by mammalian embryos are examples of how organisms construct their own developmental environment. The female goldenrod gallfly lays its eggs on the goldenrod. When they hatch, the caterpillars eat the goldenrod stem, and their salivary proteins induce cell proliferation in the goldenrod, triggering formation of a gall. The larva enters the gall, which becomes its home, and continues eating from within. As winter approaches, the larva begins to produce sugars that act as an antifreeze. The trigger for this synthesis is not temperature but aromatic substances produced by the desiccating gall (Williams & Lee, 2005). Here, we see both reciprocal induction on ecological level and reciprocal causation at evolutionary level. The gallfly larva creates a niche by causing the plant to change its development, and that niche provides safety, nutrition, and the cue for the larva to change its development as winter approaches.
Gallfly larvae create a home for themselves by inducing plants to produce galls.
Mammalian development provides another example. Mammalian embryos construct their niche by instructing the uterus to alter its cell cycles, its adhesion proteins, and by inducing the formation of blood vessels. The fetus induces changes in the uterus preparing it for implantation of an embryo, thereby causing the uterus to become a habitat for the developing organism. In return, the uterus reciprocally helps induce the formation of the placental tissues of the embryo (Spencer et al. 2004; Ticconi et al. 2006).
Niche construction during development can also be the product of behavior. For instance, female dung beetles manufacture and bury a brood ball of dung and insert into it a faecal pedestal onto which they lay an egg. Through this niche construction they provide a safe home, food supply and microbiome for their developing young (Schwab et al. 2016). The developing larvae also engages in niche construction, in a manner that affects its development. It processes the brood ball in ways that propagate and change the composition of the microbiome. Experiments show that both maternal and offspring niche construction strongly affect offspring size, fitness and trait characteristics, such as sexual dimorphism (Schwab et al. 2016, 2017). This example illustrates how niche construction can both shape developmental trajectories and affect evolutionary outcomes. Ongoing work is exploring how beetle populations diverge in their reliance on certain forms of niche construction, and how it affects range expansions and reproductive isolation.
Organisms construct their environment and alter each other
One of the most exciting areas of developmental niche construction concerns symbiotic bacteria. Here, the bacteria are both part of the environment and an organism seeking a niche. Bacteria and developing animal are environments for each other, and scaffold each other’s development (Chiu and Gilbert 2015). Bacteria are an essential component of the inheritance system in many insects and vertebrates. For instance, Dedeine et al. (2001) found that females of the wasp Asobara tabida cannot make their oocytes without products being made from the Wolbachia bacteria stored in them: A. tabida treated with antibiotics were unable to produce mature eggs.
Similarly, mammalian development is not complete without signals from symbiotic bacteria (Hooper et al. 2001; Xu and Gordon 2003). Mice bred without gut bacteria have aberrant digestive systems and defective immune systems. The bacteria induce gene expression in intestinal epithelia, and these genes are responsible for activating the pathways that allow intestinal capillaries to form and lipids to be transported (Hooper et al. 2001; Stappenbeck et al. 2002). Without these microbes, mice lack the capillary vasculature of the intestinal villi.
Ley et al. (2006) have shown that human babies acquire their gut microbial communities from the vagina and the feces of their mothers early in life. Babies born through Caesarian section had an altered colonization pattern compared with vaginally delivered babies.
Niche construction by symbionts has been studied extensively in the squid Euprymna scolopes and the luminescent bacterium Vibrio fischeri (McFall‐Ngai & Ruby, 1991; Montgomery & McFall‐Ngai, 1994). The adult squid is equipped with a light organ composed of sacs filled with light‐emitting bacteria, but newly hatched squid have neither the bacteria, nor the light organ to house them. Rather, the symbiotic bacteria interact with the larval squid to build their niche. The juvenile squid acquires the bacteria from seawater by pumping through its mantle cavity (Nyholm et al. 2000). The bacteria bind to a ciliated epithelium in this cavity, and secrete chemicals that induce hundreds of genes in the epithelium to become active, leading to the differentiation of the surrounding cells into storage sacs for the bacteria, and the expression of genes encoding opsins and other visual proteins in the light organ (Chun et al. 2008; Koropatnick et al. 2004; McFall‐Ngai. 2008; Tong et al. 2009). In this mutualistic arrangement, both organisms change their gene expression patterns, in a beneficial way. The bacteria get a niche, and the squid develops a light organ that allows it to swim at night.
Symbiotic bacteria in the squid gut induce the squid’s genes to produce a light organ.
Another interesting case of bacterial niche construction concerns the formation of the rumen of ruminants such as cattle, sheep and deer. Newborn calves have sterile rumens, and the digestive tube becomes colonized by microbes as the calf pass through the birth canal. When the calf starts eating solid food, bacteria produce plant-wall‐digesting enzymes that metabolize the polysaccharides. Over 70% of the cow’s energy comes from this microbial digestion of plant fiber (La Reau & Suen, 2018). The bacteria in the rumen multiply when given this food, and as they proliferate, they produce volatile fatty acids, that cause the rumen to grow and complexify (Gilbert, 2020). Here, the gut bacteria help construct their niche, the rumen, without which the cattle could not survive.
Many experiments now show that symbionts are not merely passive travelers, but provide essential functions for their hosts. In mammals, it has been estimated that 50-90% of cells are symbionts (Bäckhed et al. 2005; Sender et al. 2016). For instance, about one-third of the metabolites in our blood are derived from bacteria, whilst symbionts have been shown to be critical for normal brain development, and effective functioning of the immune system (Gilbert & Epel 2009; Gilbert et al. 2012). The bacteria of the gut are critical in the organogenesis of the gut capillaries and lymphoid tissues (Round et al. 2010).
Microbiomes (symbionts such as bacteria, archaea, protists, fungi and viruses) can be transmitted from one generation to the next, including via eggs and seeds, through the birth canal in mammals, or by drinking mother’s milk (Gilbert et al. 2012; Roughgarden et al. 2018). Many organisms rely on their microbiome to carry out essential processes: for example, corals rely on microalgae for photosynthesis and energy production, termites rely on intestinal protists to digest cellulose, and legumes require rhizobial bacteria for nitrogen fixation (Kamra 2005; Gilbert et al. 2012; Roughgarden et al. 2018).
“Development is a multi-species project” (Chiu & Gilbert 2015), and microbial communities do not merely “occupy” hosts but rather host and microbiome “are constantly constructing and modifying each other as local niches” (Chiu & Gilbert 2015).
Extra-genetic inheritance and the ‘start-up niche’.
The inheritance of symbionts from the mother is an example of extra-genetic inheritance. Recent years have witnessed the accumulation of empirical support for extra-genetic inheritance through multiple pathways, including epigenetic, ecological, behavioral, and cultural inheritance systems and the transmitted microbiome (Bird 2002; Jablonka & Lamb 2005; Bonduriansky & Day 2018). Diverse resources are transmitted from parents to offspring, including components of the egg, hormones, symbionts, epigenetic marks, prions, small RNAs, antibodies, ecological resources and learned knowledge.
Developmental pathways known to affect trait inheritance
Traditionally considered proximate causes of development, it is now evident that some of these factors can lead to the inheritance of phenotypes, and many researchers now attempt to integrate these components into an extended concept of heredity (Bonduriansky & Day 2018; Danchin et al. 2011; Jablonka & Lamb 2014). These additional pathways allow for ‘soft’ inheritance (environmental influences on heredity). Recognition of extra-genetic inheritance has important implications, both for understanding development and comprehending how development can influence evolution.
Offspring inherit not just genes but a ‘start-up niche’, comprising a specific parentally chosen location for birth and a transmitted package of resources (Odling-Smee 2010). For example, phytophagous insects typically choose to lay eggs on specific host plants, which become food for their larvae, whilst in birds, nutrients and hormones are provided in the yolk for embryonic nutrition. Many organisms provide protective chemicals for their offspring in this start-up niche, including antibodies such that the young can survive before their immune systems mature (birds, mammals), compounds that are poisonous or distasteful to predators (moths) (Dussourd et al. 1988), or even sun-blocks that protect transparent embryos and larvae from the effects of solar radiation (Goldstone et al. 2006).
From the niche-construction perspective, the key task for any developing organism becomes the active regulation of its inherited ‘niche’, by responding to its environment, and by altering its environment, in ways that keep its personal organism–environment relationship continuously adaptive, for the rest of its life. This active ‘niche regulation’ undermines the (generally assumed) independence of environments from developing organisms: just like selective environments, developmental environments are not independent on the organism. Niche construction provides a pathway through which developmental processes can influence evolutionary processes.
FJ, Gilbert SF. 2008. EvoDevo and niche construction: building bridges. Journal of Experimental Zoology Part B. 310:549–566. Points to common ground between niche construction and evo devo. Argues that the same conceptual barriers have hindered both.
Human niche-constructing activities may inadvertently promote disease. Cultural practices, such as cultivating crops and keeping animals, appear to have aided the spread of diseases, such as malaria and sickle-cell anaemia, leading to the selection of alleles that confer resistance to the promoted diseases (Durham 1991; Laland et al. 2010; O’Brien & Laland 2012).
Many human genes that provide some immunity from, or resistance to, disease or pathogens are now known to have been subject to recent selection, and are thought to have been promoted by agriculture or other farming practices (Laland et al. 2010).
The inadvertent construction of disease niches by human cultural activities may have been going on for some time, and is implicated in the emergence of tuberculosis, an ancient human disease. The controlled use of fire may have triggered the spread of TB, through a combination of increased opportunities for transmission brought about by the developing social culture that fire use encouraged and the lung damage that smoke inhalation caused (Chisolm et al. 2016).
The control of fire may have led to the spread of tuberculosis amongst ancestral humans.
Another system that exhibits feedback between human cultural niche construction and genetic selection is the host-parasite relationship between antibiotic treatment and viability selection for antibiotic-resistant bacterial strains. This is an example of inter-specific cultural niche construction. Boni and Feldman (2005) found that the cultural transmission of antibiotic use favours selection of resistant bacterial strains, which in turn can result in cultural selection for the avoidance of antibiotic use.
This kind of host behaviour can result in the classic niche-construction phenomenon of maintaining strain polymorphism even in parameter regions where it would not otherwise be expected. Interestingly, the evolution of either the host activity or the parasite strain can be viewed as a niche-constructive activity that modifies the selective environment of the other. Potentially this promotes an arms race between two types of transmitted information, cultural and genetic.
Here, the niche constructive effects can be described fully in terms of trait–trait co-evolution as there is no ecological inheritance of a constructed ‘resource’ that is separate from the cultural or the genetic information transmission systems. The relative frequencies of bacterial resistance and sensitivity are the effective ‘resource’ influencing the cultural evolution of antibiotic treatment, and visa versa.
Disease vectors too can construct niches. For instance, as obligate parasites, viruses too are constantly modifying the host environment, and these modifications are now thought to drive evolutionary feedback between the virus and its host, across multiple scales, from cells to ecosystems (Hamblin et al. 2014). For instance, the rabies virus causes aggression of the host and thereby facilitates its own propagation.
Hamblin SR, White PA, Tanaka MM. 2014. Viral niche construction alters hosts and ecosystems at multiple scales.Trends in Ecology & Evolution.29(11): 594-9. Viruses modify host environments, and these modifications drive evolutionary feedback between the virus and its environment across multiple scales from cells to ecosystems.
Niche construction and conservation biology
The impact of niche construction on diversity
Organisms do considerably more in ecosystems than compete with and eat each other (trophic interactions). They also produce, modify, and destroy habitat and resources, in the process driving co-evolution and regulating hydrological, nutrient, and element (e.g., carbon) cycling (Odling-Smee et al. 2003). Through niche construction and ecosystem engineering, organisms create ‘engineering’ control webs that affect the stability and productivity of ecosystems (Jones et al. 1994, 1997).
Across scales that encompass both the presence and absence of ecosystem engineering/niche construction, the net effect should be to enhance species richness via a net increase in habitat diversity (Jones et al. 1997). Recent studies provide support for this hypothesis. For example, natural sites with and without beavers exhibit low overlap in species composition. By increasing habitat heterogeneity, beavers increased herbaceous plant species numbers by more than 33% (Wright et al. 2002).
Another example is seaside arrowgrass (Triglochin maritima), which facilitates plant diversity in salt marshes by creating elevated rings (maintained structurally by its rhizomes) with increased reductive potentials and reduced salinity. T. maritima supports both a greater abundance of species and the growth of species not present in the adjacent substratum (Fogel et al. 2004).
Brathen & Raivolainen (2015) show that in tundra plant communities, forbs and grasses were the least abundant growth forms, yet they had a strong positive effect on species diversity through their effects on ecosystem process rates and nutrient cycling. The effects of these plants are completely disproportionate to their share of biomass.
Niche construction and conservation strategies
Findings like these have important implications for understanding, managing, and conserving ecosystems (Crain & Bertness 2006; Boogert et al. 2006; Laland & Boogert 2010). Academics and politicians alike have failed to consider fully both the important role that these control webs play in ecosystems, and the consequences of human activities that destroy those webs of connectance.
If ecosystems are threaded by ‘engineering’ control webs, then the disappearance of key niche constructors may lead to abrupt changes in the resources and selection created by them, greatly affecting other species. Populations that have become dependent on engineered habitat and resources may be unable to cope with the loss, while genetic adaptation across generations is often too slow to counteract environmental modifications, leading to further declines in biodiversity and ecosystem functioning. This highlights the importance of preserving species that construct or maintain habitat and resources for other species (Crain & Bertness 2006; Boogert et al. 2006; Ehlers et al. 2008).
One might avoid unforeseen negative consequences of introducing engineering species by replenishing the niche constructors’ effects on the environment, rather than the organisms themselves (Odling-Smee et al. 2003). Some examples of the mimicking of engineering effects include introduction of artificial mussel mats (Crooks & Khim 1999), artificially created leaf ties otherwise produced by caterpillars (Lill & Marquis 2003), and relocalization of natural structures to provide lizard refuges (Pringle 2008). Research is required to explore the extent to which these manipulations can be successful, affordable and feasible at scales relevant to conservation goals.
Boogert et al. (2006) suggest an implementation strategy to conserve ecosystems through the conservation of niche-constructing activities. The strategy includes the following steps:
Step 1) Set conservation goals for the target ecosystem.
Step 2) Determine the key engineers in the target ecosystem, which will often require additional research.
Step 3) Conduct pilot studies to explore which of the following procedures might be most feasible and effective: (a) enhancing key engineers’ current activity by introducing more conspecifics or providing them with the resources required for population growth, (b) enhancing key engineering activities by introducing different species that engineer in the same manner, (c) introducing artificially manufactured products of the key engineers, and (d) creating optimal levels of abiotic and biotic factors to facilitate key engineers through trophic or nontrophic links. When the ecosystem is negatively affected by invading engineers, one could investigate the effectiveness of equivalent steps to reduce their impact.
Step 4) Implement the optimal engineering strategy or combination of strategies on a small scale. Follow up by monitoring and assessment.
Step 5) If step 4) has the desired outcome, implement the successful engineering strategy on a large scale.
Humans have dramatically altered their niche through farming and agricultural practices.
Niche construction and the Anthropocene
Unlike any other species in Earth’s history, human niche construction is so potent that we have gained the capacity to transform the functioning of an entire planet (Ellis 2015; Waters et al. 2016).
Though contemporary rates and scales of anthropogenic environmental change are unprecedented, human societies began transforming Earth’s ecology many thousands of years ago (Smith & Zeder 2013; Boivin et al. 2016).
Anthroecology theory holds that human societies scaled up and gained the capacity to transform Earth’s functioning through a long-term evolutionary process, in which increasing societal scales and increasingly transformative ecosystem engineering are coupled through positive feedbacks (known as ‘runaway sociocultural niche construction’) (Ellis 2015, Ellis 2018).
Runaway cultural niche construction is transforming planet Earth.
Much attention has been given to how human activities can have a negative impact on biodiversity, and rightly so, but it is important to recognize that human impacts have multitudinous, complex and often conflicting effects on the biosphere. Counter-intuitively, recent research shows that human niche construction is actually increasing biodiversity in restricted contexts. For instance, it is known that a complex ecosystem of attached organisms develops on submerged structures (such as oil rigs) in the marine environment, which supports a localized food web that could not exist without them (Fortune & Paterson, 2018). The impact of human activity on the environment needs to be assessed on a case-by-case basis, drawing on careful science.
Complex ecosystems can develop around submerged structures in marine environments. Fig 1 of Fortune & Paterson, 2020.
Jones CG, Lawton JH, Shachak M. 1994. Organisms as ecosystem engineers.Oikos. 69: 373-386. The most authoritative introduction to the concept of ecosystem engineering.
We use cookies to optimize our website and our service.
Functional
Always active
The technical storage or access is strictly necessary for the legitimate purpose of enabling the use of a specific service explicitly requested by the subscriber or user, or for the sole purpose of carrying out the transmission of a communication over an electronic communications network.
Preferences
The technical storage or access is necessary for the legitimate purpose of storing preferences that are not requested by the subscriber or user.
Statistics
The technical storage or access that is used exclusively for statistical purposes.The technical storage or access that is used exclusively for anonymous statistical purposes. Without a subpoena, voluntary compliance on the part of your Internet Service Provider, or additional records from a third party, information stored or retrieved for this purpose alone cannot usually be used to identify you.
Marketing
The technical storage or access is required to create user profiles to send advertising, or to track the user on a website or across several websites for similar marketing purposes.












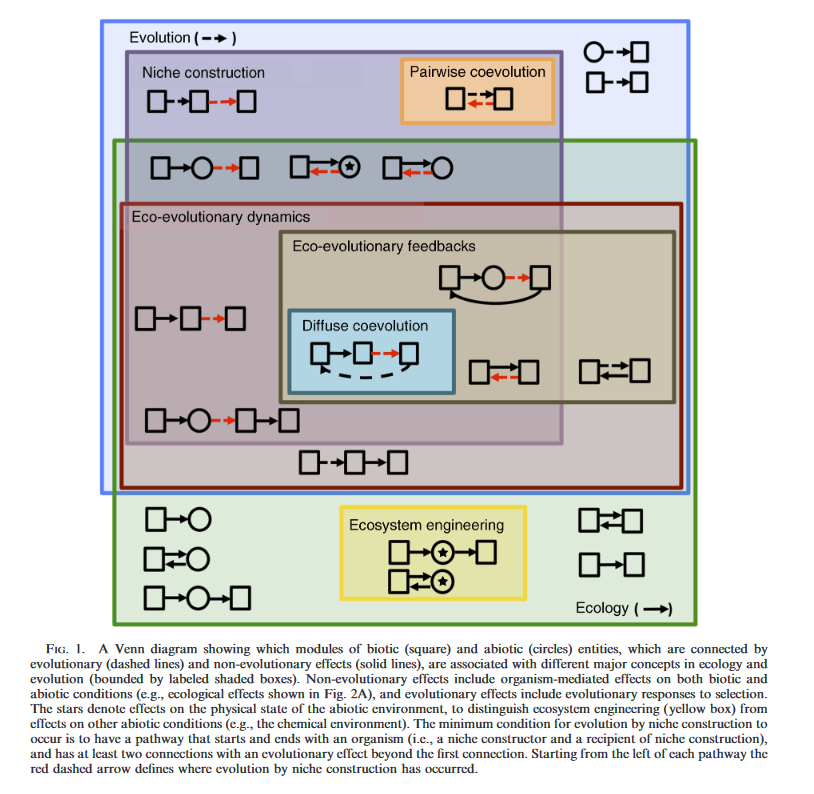



 Blog article: “Is social transmission a forgotten force in coevolution?” by Rose Thorogood
Blog article: “Is social transmission a forgotten force in coevolution?” by Rose Thorogood


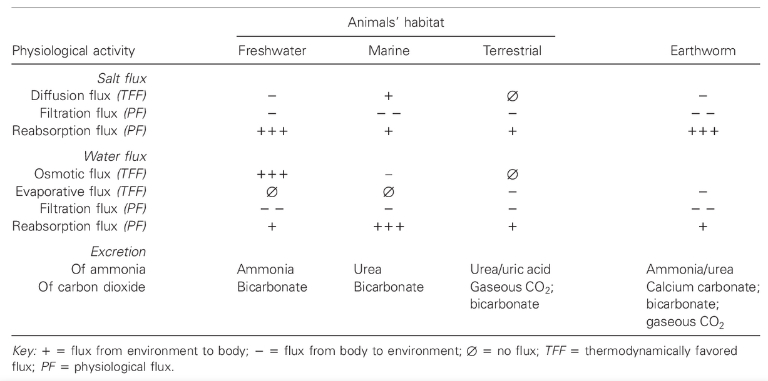
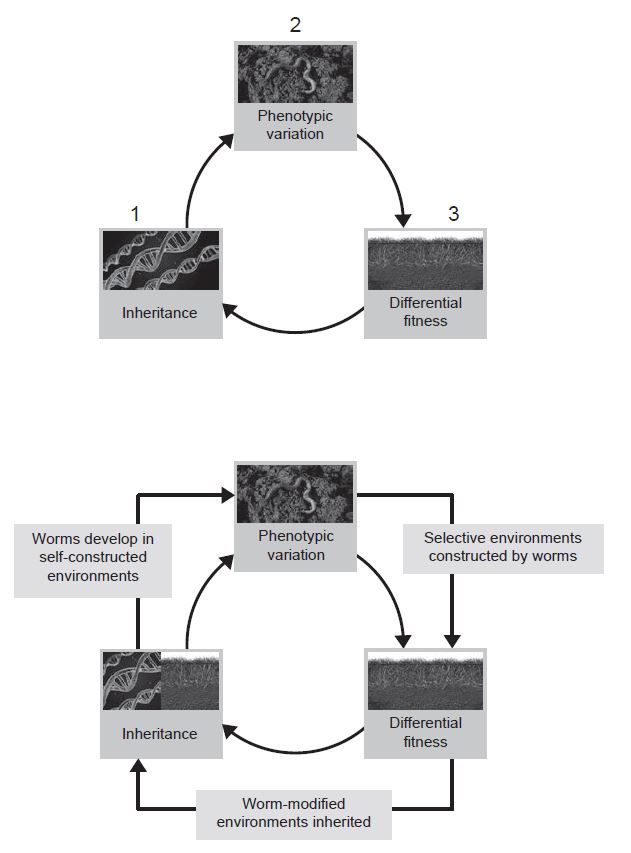

 Mayr’s account of causation remains widely accepted today, with both positive and negative ramifications. One negative consequence was that Mayr insisted on a strict separation of proximate and ultimate causes:
Mayr’s account of causation remains widely accepted today, with both positive and negative ramifications. One negative consequence was that Mayr insisted on a strict separation of proximate and ultimate causes: